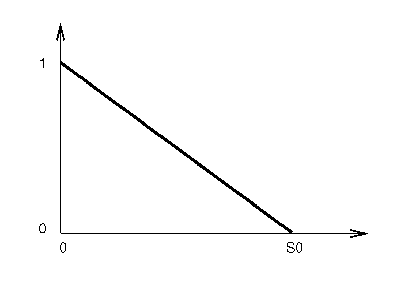Les algorithmes évolutionnaires sont des algorithmes d’optimisation s’appuyant sur des techniques dérivées de la génétique et de l’évolution naturelle1 : croisements, mutations…
Encore peu connus en France2, leur popularité est croissante aux États-Unis surtout depuis 1985.
Ce sont des méthodes d’optimisation stochastique qui ne requièrent a priori pas de régularité sur les fonctions optimisées : les applications possibles sont très diverses. Ils ont la propriété de pouvoir localiser plusieurs optima mais présentent deux inconvénients majeurs. D’une part, ce sont des méthodes chères en calculs qui ne garantissent pas l’optimalité du résultat et l’on conseille généralement d’appliquer ce type de méthodes lorsque rien d’autre ne marche. D’autre part, la mise en oeuvre pratique de ces méthodes demande une grande expérience et les résultats théoriques sont généralement peu utiles en pratique.
On distingue quatre familles historiques :
Un algorithme génétique réalise une optimisation dans un espace de données. Pour pouvoir utiliser les techniques génétiques il faut disposer de deux opérations :
Ces deux éléments sont les seuls éléments spécifiques au problème à résoudre. Une fois qu’ils sont fixés, l’algorithme génétique que l’on appliquera sera toujours le même, que l’on optimise la distribution de gaz dans un pipeline, la trajectoire de deux mobiles qui doivent s’éviter, ou la fonction énergie d’un réseau de neurones.
Il s’exécutera en plusieurs étapes :
Nous allons voir, dans le détail et sur un exemple, comment fonctionne un algorithme génétique.
Nous allons considérer l’exemple suivant : soit la fonction3 f(x)= 4x(1−x) prenant ses valeurs sur [0,1[. Nous souhaitons trouver le maximum de cette fonction.
Pour coder les données, on peut se contenter de multiplier x par 28 et de prendre la partie entière du résultat. On obtient ainsi un nombre entier appartenant à l’intervalle [0,28−1]. Sa représentation en binaire donne une séquence de 8 bits4 le codant. Ainsi x=0.3 multiplié par 28 donne 76.8, dont on prend la partie entière : 76. 76 en binaire donne : 01001100. C’est la séquence de bits codant 0.3.
La fonction U(x) est ici : U(x)=f(x). Il s’agit donc exactement de la fonction à maximiser.
Les deux fonctions étant fixées, il ne reste qu’à appliquer l’algorithme génétique.
Première étape : nous commençons par générer aléatoirement les séquences qui vont composer notre population. Nous ne générons que 4 séquences pour cet exemple particulier5. Nous présentons dans le tableau 1.1 la population initiale générée, ainsi que la valeur de la fonction d’adaptation et le pourcentage de reproduction pour chaque séquence dans les trois premières colonnes.
Séquence Valeur U(x) % de chance % cumulés Après de reproduction reproduction 10111010 0.7265625 0.794678 0.794678 / 2.595947 = 0.31 0.31 11011110 11011110 0.8671875 0.460693 0.460693 / 2.595947 = 0.18 0.31+0.18=0.49 10111010 00011010 0.1015625 0.364990 0.364990 / 2.595947 = 0.14 0.49+0.14=0.63 01101100 01101100 0.4218750 0.975586 0.975586 / 2.595947 = 0.37 0.62+0.37=1.00 01101100 = 2.595947
Nous devons ensuite effectuer la deuxième étape, la reproduction. Pour savoir quels sont les quatre éléments de la population suivante avant croisement, on tire aléatoirement quatre nombres entre 0 et 1. La comparaison du nombre avec le pourcentage cumulé de la quatrième colonne du tableau 1.1 permet de savoir quel est l’élément généré. Si nous tirons les 4 nombres 0.47, 0.18, 0.89, 0.75, nous obtenons la colonne 6 du tableau comme population après reproduction.
Passons maintenant au croisement. Un certain nombre de séquences de la population vont être remplacées par des croisements avec d’autres membres de cette population. Typiquement, de 25 à 75% des séquences sont laissées intactes, les autres étant remplacées par des croisements. Supposons que le sort désigne les séquences 1 et 3 comme devant être croisées et remplacées par les résultats du croisement. Il faut encore tirer aléatoirement un nombre entre 1 et 7 qui va désigner l’endroit où le croisement va s’effectuer ; supposons que ce soit 5. Le croisement est alors représenté sur la figure 1.1.
La population après croisement est représentée dans le tableau 1.2.
Après croisement Valeur U(x) 01101110 0.4296875 0.980225 10111010 0.7265625 0.794678 11011100 0.8593750 0.483398 01101100 0.4218750 0.975586
Il resterait à appliquer les mutations. Mais le pourcentage de mutation est toujours très faible (inférieur ou très inférieur à 10%) et nous supposons qu’il n’y en a pas ici.
Nous constatons que non seulement la moyenne des valeurs d’adaptation est meilleure, mais que la qualité de la meilleure séquence est plus grande.
Comme souvent pour ce type de mécanisme qui pourrait apparaître comme un peu << magique >> (les réseaux de neurones font partie de cette catégorie), les difficultés sont invisibles, mais bien réelles. La première d’entre elles consiste à bien choisir les divers paramètres : pourcentage de croisement, pourcentage de mutation, taille de la population…
Mais le problème le plus important est le codage des données. Comme le dit Goldberg lui-même, le codage des données est un art, et de cet art dépend le succès ou l’échec de la tentative. Dans un cas aussi simple que celui-ci, le codage parait évident et pourtant, on remarquera que les séquences 10000000 et 01111111 sont très bien adaptées l’une et l’autre, bien que leur codage soit fondamentalement différent, ce qui n’est pas une bonne propriété6. La théorie des graphes présentée dans la section 1.3.2 permet de comprendre ce qu’est un bon codage de données.
Plusieurs approches ont été envisagées. La théorie des schémas présentée dans la section 1.3.2 est la plus simple à aborder et donne dans un cadre simplifié une idée du mécanisme de convergence des AGs. On trouve néanmoins plusieurs démonstrations de convergence théorique des algorithmes évolutionnaires dont :
Nous allons d’abord poser quelques définitions : [Séquence] On appelle séquence A de longueur l(A) une suite A=a1a2… al avec ∀ i∈ [1,l], ai ∈ V={0,1}. Ceci correspond à la notion de séquence de bits que nous avons utilisée jusqu’à présent.
Nous avons également besoin de la notion de schéma et de quelques définitions associées : [Schéma] On appelle schéma H de longueur l une suite H=a1a2… al avec ∀ i∈ [1,l], ai ∈ V+={0,1,*}. Comme nous allons le voir, une * en position i signifie que ai peut être indifféremment un 0 ou un 1.
[Instance] On dit qu’une séquence A=a1… al est une instance d’un schéma H=b1… bl si pour tout i tel que bi≠ * on a ai=bi. Ainsi, H=010*0101 est un schéma et les séquences 01000101 et 01010101 sont des instances de H (ce sont même les seules).
[Position fixe, position libre] Soit un schéma H. On dit que i est une position fixe de H si ai=1 ou ai=0. On dira que i est une position libre de H si ai= *.
[Ordre d’un schéma]
On appelle ordre du schéma H le nombre de positions fixes de H. On note l’ordre de H o(H). Par exemple, le schéma H=01**10*1 a pour ordre o(H)=5, le schéma H′=*****101 a pour ordre o(H′)=3. On remarquera qu’un schéma H de longueur l(H) et d’ordre o(H) admet 2(l(H)−o(H)) instances différentes.
[Longueur fondamentale] On appelle longueur fondamentale du schéma H la distance séparant la première position fixe de H de la dernière position fixe de H. On note cette longueur fondamentale δ(H). Ainsi, le schéma H=1**01*** a pour longueur fondamentale δ(H)=5−1=4, le schéma H′=1******1 a pour longueur fondamentale δ (H′) = 8−1=7, et pour le schéma H >>=**1***** nous avons δ(H >>)=3−3=0.
[Adaptation d’une séquence] On appelle adaptation d’une séquence A une valeur positive que nous noterons f(A). f est ce que nous avions appelé jusqu’à présent la fonction d’utilité7.
[Adaptation d’un schéma] On appelle adaptation d’un schéma H la valeur
| f(H) = |
|
où les Ai décrivent l’ensemble des instances de H. L’adaptation d’un schéma est donc la moyenne des adaptations de ses instances.
Soit un ensemble S={A1,…,Ai,…,An} de n séquences de bits tirées aléatoirement. Durant la reproduction, chaque séquence Ai se reproduira avec une probabilité :
| pi= |
|
Supposons que nous ayons à l’instant t un nombre m(H,t) de séquences représentant le schéma H dans la population S. À l’instant t+1 nous devons statistiquement avoir un nombre :
| m(H,t+1)=m(H,t) . n . |
|
Posons
| = |
|
ft représente la moyenne de l’adaptation des séquences à l’instant t. La formule précédente devient :
| m(H,t+1)=m(H,t) |
|
Posons ct(H)=f(H)/ft−1. On obtient alors :
| m(H,t+1)=(1+ct(H)) m(H,t) |
On voit donc qu’un schéma, dont l’adaptation est au-dessus de la moyenne, voit son nombre de représentants augmenter, suivant une progression qui est de type géométrique si nous faisons l’approximation que ct(H) est constant dans le temps. Nous avons alors :
| m(H,t)= (1+c(H))t . m(H,0) |
Le résultat est donc clair : si la reproduction seule était en jeu, les schémas forts élimineraient très rapidement les schémas faibles.
Nous allons nous intéresser à la probabilité de survie ps(H) d’un schéma H lors d’une opération de croisement. Considérons cela sur un exemple. Soit le schéma H=**10*1**. Supposons qu’une séquence qui est une instance de ce schéma soit croisée avec une autre séquence. Quelle est la probabilité pour que la séquence résultante soit encore une instance de H ? Il est impossible de répondre exactement à la question, tout au plus peut-on donner une borne inférieure de cette valeur. Il est clair que H ne sera pas détruit si le site de croisement qui est tiré au sort est inférieur à 3 (avant le premier 1) ou s’il est supérieur à 5 (après le dernier 1)8.
On voit donc immédiatement qu’une borne inférieure de la probabilité de détruire un schéma H est δ(H)/(l−1). Donc la probabilité de survie dans un croisement est 1−δ(H)/(l−1). Si d’autre part on ne croise qu’une fraction pc de séquences dans une population donnée, la probabilité de survie est donnée par :
| ps≥ 1−pc |
|
En rassemblant ce résultat et celui du paragraphe précédent, nous obtenons pour l’évolution d’une population9 :
| m(H,t+1) ≥ m(H,t) (1+ct(H)) (1−pc |
| ) |
Supposons que la probabilité de mutation d’un bit dans une séquence soit pm. Dans un schéma H, seules les positions fixes peuvent être détruites. Comme la probabilité de survie d’un bit est 1−pm, la probabilité de survie d’un schéma H contenant o(H) positions fixes est (1−pm)o(H). La probabilité de mutation étant toujours petite devant 1, on peut faire un développement limité au premier ordre, ce qui nous donne une probabilité de survie de 1−o(H)pm.
En introduisant ce terme, nous avons donc l’équation finale :
| m(H,t+1) ≥ m(H,t) (1+ct(H)) (1−pc |
| −o(H)pm) |
La formule ci-dessus nous apprend deux choses :
Ceci enseigne une chose fondamentale pour le codage de données : les schémas qui codent les données << intéressantes >> pour le problème doivent avoir un ordre et une longueur fondamentale faibles10, alors que les données << sans intérêt >> doivent être codées par des schémas qui ont un ordre et une longueur fondamentale élevés.
Les résultats présentés ci-dessus ne sont qu’une introduction à la théorie des schémas, il existe de nombreux approfondissements. On trouvera dans les références suivantes les principaux modèles théoriques sur la théorie des schémas et ses extensions :[Gol89b, Vos91, BM93, Gol89a, BG87, CS88, Moo94, DM92, SGE91]
Nous allons tester l’utilisation des algorithmes génétiques sur un cas très simplifié d’optimisation de trajectoires d’avions dans le plan horizontal. Pour plus détails, on peut se reporter à [Dur96]. Nous posons comme hypothèses : algorithmes!génétiques!exemple complet
Un avion est ici défini comme un point matériel muni d’un vecteur vitesse (dans l’exemple qui suit, les avions volent à 500 nœuds ou 900 km/heure). Nous nous sommes limités au plan horizontal car, en contrôle aérien, on préfère donner aux avions des manœuvres dans le plan horizontale plutôt que des manœuvres de montée ou de descente (leur coût en termes de consommation et d’exécution est plus faible, le confort du passager est préservé). Le temps est discrétisé par pas de 10 secondes.
Le codage binaire présenté dans la section 1.2.2 pourrait être utilisé dans le cas présent, mais on lui préférera un codage plus immédiat qui préserve la structure des données. La manœuvre effectuée par un avion est déterminée par un triplet (a, t0, t1) où a∈ {−30,−20,−10,0,10,20,30}, t0 et t1 sont des variables réelles comprises entre 0 et 20 minutes. C’est ce triplet qui sera retenu comme codage, les opérateurs de croisement et de mutation devront être adaptés en conséquence. Un problème à n avions sera codé par un 3 n -uplet.
Le critère d’évaluation retenu s’exprime de la façon empirique suivante:
|
où C est la durée totale de conflits (somme des durées des périodes pendant lesquelles deux avions sont à moins de 15 km l’un de l’autre), N est le nombre de manœuvres (on considère qu’il y a manœuvre si a≠ 0 et t0<t1) et R est le retard moyen à l’arrivée relatif à la durée du vol (20 minutes). D’autres choix sont possibles, celui-ci garantie qu’une solution sans conflit (>1/2) est toujours plus adaptée qu’une solution avec conflit (<1/2). Par ailleurs, N est entier et R<1 donc le critère d’évaluation minimise le nombre de manœuvres avant de minimiser le retard.
Compte-tenu du codage des données choisi, nous devons définir les opérateurs de croisement et de mutation.
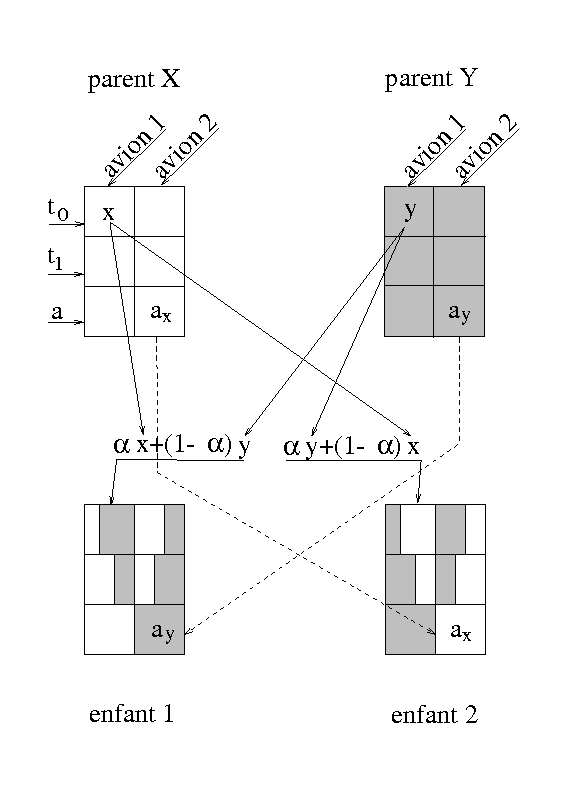
L’opérateur de croisement choisi (voir figure 1.2) est le suivant :
L’opérateur de mutation choisi aléatoirement avec des probabilités identiques la variable t0, t1 ou a de l’avion 1 ou 2 d’un élément de population et le modifie en lui ajoutant un bruit gaussien (s’il s’agit de t0 ou t1) ou en lui ajoutant ou retranchant 10 degrés selon les possibilités (s’il s’agit de a).
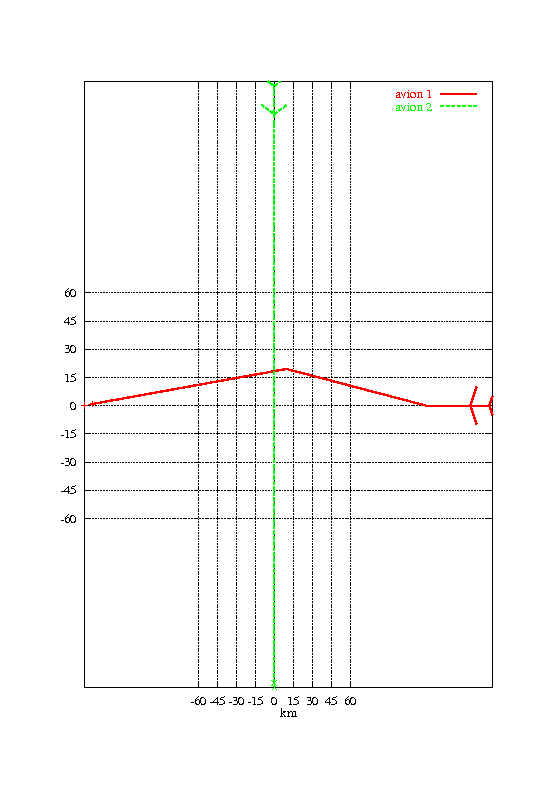
La figure 1.3 montre une solution obtenue après 28 générations de l’algorithme génétique. Les paramètres choisis étaient :
La durée d’une génération est alors de l’ordre de 3 centièmes de seconde sur un Pentium II 450. L’optimisation complète dure moins d’une seconde.
Cette solution est en fait optimale (ce résultat ne pouvant être assuré a priori, mais seulement constaté a posteriori). Un algorithme de type A*, appliqué au problème, fournit un résultat équivalent en un temps équivalent. L’algorithme génétique est donc, pour cet exemple, équivalent à une méthode de parcours de graphe.
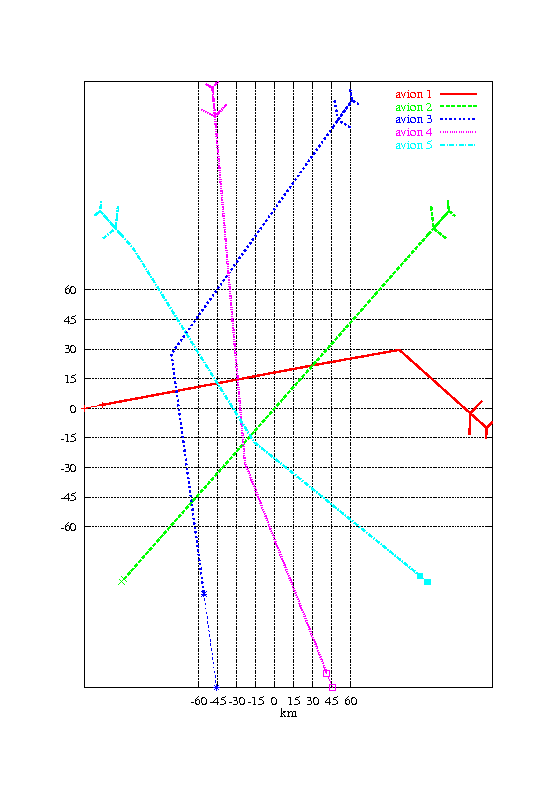
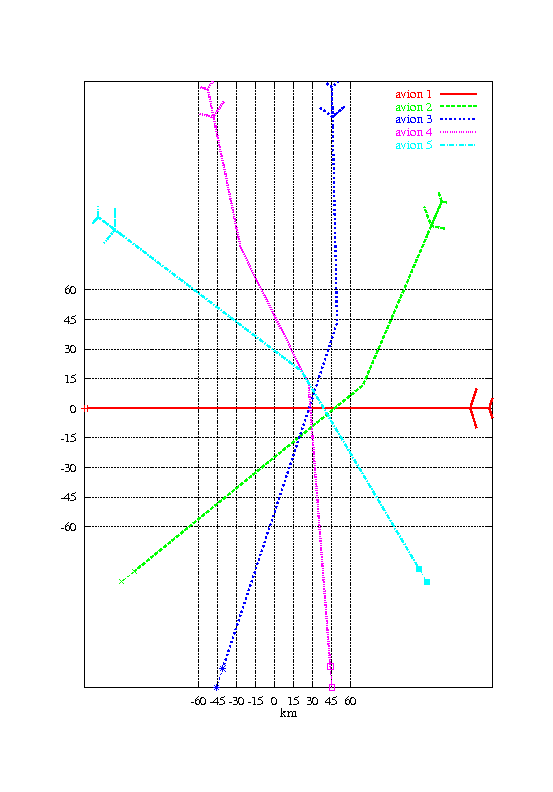
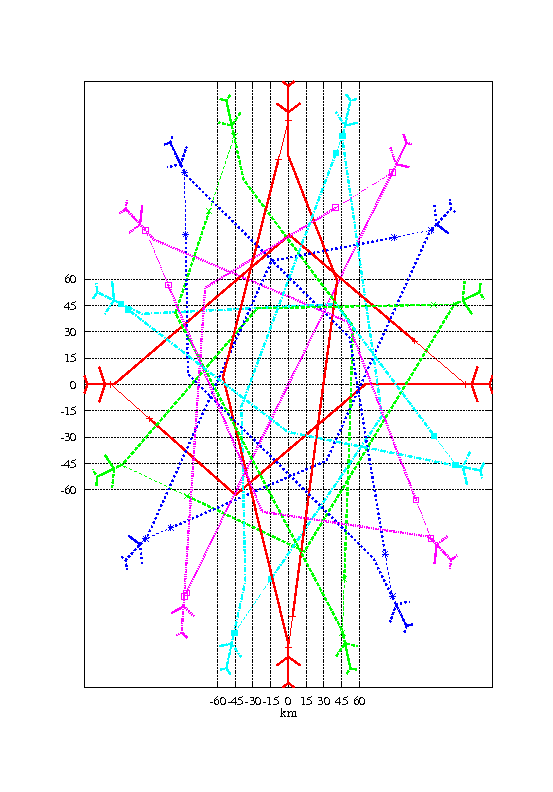
Nous avons ensuite voulu tester l’algorithme dans le cas où le conflit implique 5 avions initialement situés régulièrement sur un demi-cercle de 150 km de rayon et convergent vers le centre du cercle. La taille de la population a été augmentée à 500 individus. Les autres paramètres sont restés inchangés.
La figure 1.4 montre une solution obtenue après 200 générations de l’algorithme génétique. Le temps de calcul est de 1mn 20s.
La solution obtenue n’est pas optimale. Nous verrons par la suite comment on peut l’améliorer en utilisant des méthodes de sharing (section 1.5.5) et la séparabilité partielle du problème (section 1.5.6).
L’algorithme A* ne permet pas de trouver la solution optimale car son temps d’exécution est trop long. 20mn de vol représentent 120 pas de temps de 10s. Un avion qui n’a pas encore engagé de manœuvre a 7 possibilités de modification de cap à chaque pas de temps (de -30 à 30 degrés par pas de 10). S’il a engagé une manœuvre, il peut soit la poursuivre, soit prendre un cap vers sa destination. Une fois sur la branche de retour, il n’a plus d’alternative. La taille de l’arbre de recherche pour chaque avion est 7+3 n (n+3) avec n=120. Si le conflit implique p avions, la taille de l’arbre devient [7+3 n (n+3)]p. Pour 2 avions, la taille reste inférieure à 1010. Pour 5 avions, on dépasse 10 23.
Nous allons examiner rapidement dans cette section différentes techniques complémentaires qui peuvent améliorer les performances des algorithmes génétiques. Ces techniques sont pour la plupart présentées en détail dans [Gol89c], à l’exception des représentations spécialisées, que l’on peut trouver dans [Mic92], avec bien d’autres.
Les techniques de codage sous forme de chaînes de bits et de croisement standard présentent un certain nombre d’inconvénients :
Il est alors naturel de tenter de définir des codages et des opérateurs spécialisés, qui s’appliqueront efficacement sur le problème considéré.
Dans l’exemple de la section 1.2.2, le codage en chaîne de bits a été abandonné au profit d’un codage utilisant directement les variables du problème. Les opérateurs de croisement et de mutations ont du alors être définis en fonction du codage. Les variables réelles et entières peuvent être traitées différemment. Ce type de représentation a pour but de mieux respecter la topologie de l’ensemble sur lequel l’optimisation est opérée à travers le codage.
Dans le cas des variables réelles, on emploie de façon classique le croisement barycentrique. Pour générer deux enfants, on commence par tirer un nombre α au hasard dans l’intervalle [−0.5,1.5] et par appliquer les formules suivantes :
|
En ce qui concerne la mutation, on génère l’élément muté en appliquant la formule :
| p1′ = p1+ B(0,σ) |
où B(0,σ) est un bruit gaussien centré en 0 et d’écart type σ. Le choix de σ dépend, bien entendu, du problème considéré.
Il est extrèmement important de réaliser que le codage et les opérateurs choisis conditionnent fortement la qualité de la solution obtenue. Les algorithmes génétiques ne sont pas des boîtes noires miraculeuses dans lesquelles il suffirait d’introduire une fonction pour en retirer l’optimum. Sans une réflexion appropriée sur le codage et les opérateurs, les résultats seront au mieux médiocres.
L’un des problèmes des algorithmes génétiques est que le sens d’un allèle est lié à sa position, position qui dépend du codage. Ainsi, supposons que dans un premier codage une propriété importante soit codée (sur 2 bits) par le premier et le huitième bit d’une chaîne de 8 bits. Avec cette forme de codage, tout croisement opérera sur cette propriété. Si au contraire la propriété était codée sur le premier et le second bit (par exemple), le bloc serait presque insensible au croisement. C’est de cette constatation que vient l’idée de réorganiser les bits dans un individu, de façon à créer des blocs stables.
Un opérateur de réorganisation opère de la façon suivante : soit l’élément de population 10100010 ; on tire au hasard deux positions, par exemple 3 et 7. Tous les bits compris entre les positions 3 et 7, c’est à dire la séquence 1000, sont alors inversés, pour donner l’individu 10000110.
Se pose alors un premier problème : dans la présentation standard des algorithmes génétiques, le codage d’une propriété dépend de la position de chaque bit. En un mot, l’individu 10100010 et l’individu 10000110 ne code plus le même objet. Il faut donc rendre le décodage indépendant de la position en stockant avec la valeur de chaque bit la valeur de sa position. Ainsi, le premier individu deviendrait (1,1),(2,0),(3,1),(4,0),(5,0),(6,0),(7,1),(8,0) et le second serait alors (1,1),(2,0),(7,1),(6,0),(5,0),(4,0),(3,1),(8,0). Dans ces conditions, les deux individus représentent bien le même objet et seront décodés de la même façon.
Il reste un dernier problème à régler : comment effectuer un croisement ? En effet, considérons les deux individus :
| (1,1),(2,0),(7,1),(6,0),(5,0),(4,0),(3,1),(8,0) |
et
| (1,1),(2,0),(3,1),(4,0),(5,0),(6,1),(7,1),(8,0) |
Supposons que le site de croisement tiré soit le milieu. Nous aurions alors les deux individus :
| (1,1),(2,0),(7,1),(6,0),(5,0),(6,1),(7,1),(8,0) |
et
| (1,1),(2,0),(3,1),(4,0),(5,0),(4,0),(3,1),(8,0) |
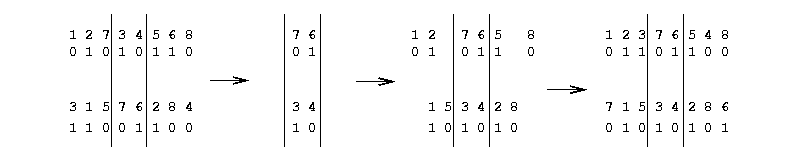
Aucun de ces individus n’est viable, puisque le premier a deux fois les positions 6 et 7 et qu’il lui manque les positions 3 et 4 et réciproquement pour le second. Il faut donc définir différemment l’opérateur de croisement. Il existe diverses stratégies (voir [Gol89c]). Nous décrirons brièvement la technique dite de partially matched crossover (PMX). Dans cette technique, on tire deux sites de croisement au lieu d’un seul (voir figure 1.5).
La zone située entre ces deux sites est d’abord échangée : on dit alors que l’allèle 3 matche (ou correspond à) l’allèle 7 et que l’allèle 4 matche l’allèle 6 (et réciproquement). Puis, on remet à leur place les allèles qui ne sont pas dupliqués après l’échange des zones de croisement. Dans le cas du premier individu, il s’agit des allèles autres que 7 et 6, et dans le cas du second, des allèles autres que 3 et 4. Puis on remplace dans l’individu du haut ce qui était l’allèle numéro 7 par son correspondant en numéro, c’est à dire l’allèle 3 de l’individu du bas ; on fait de même pour l’allèle 6, et on procède de la même façon pour les allèle 3 et 4 de l’individu du bas.
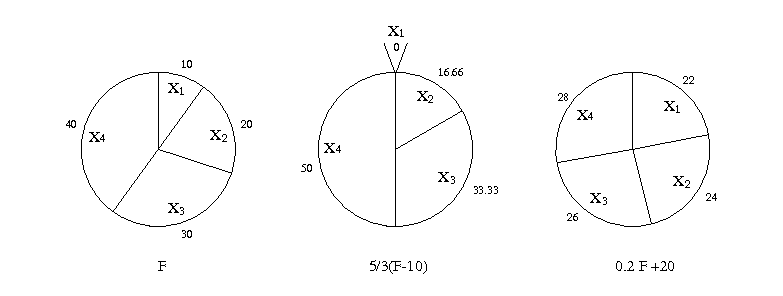
Le processus de sélection des individus qui consiste à reproduire en plus grand nombre les individus bien adaptés que les individus mal adaptés conditionne fortement la convergence de l’algorithme. La technique de mise à l’échelle ou scaling a pour but de modifier la pression de sélection. Elle consiste à composer la fonction d’évaluation avec une fonction croissante. Considérons l’exemple suivant où les 4 individus X1, X2, X3 et X4 de la population ont pour adaptation 10, 20, 30 et 40. Si l’on applique les fonctions linéaires croissantes f→ 5/3(f−10) ou f→ 0.2 f+20 sur la valeur de l’adaptation, on donne plus ou moins d’importance aux individus (voir figure 1.6). Dans le premier cas, on augmente la pression de sélection, dans le deuxième cas, on la diminue.
Le mode de sélection des individus présenté dans la section 1.2.2 se contente de reproduire les individus proportionnellement à leur critère d’adaptation. D’autres modes de sélection sont envisageables, comme la sélection par le rang (les individus sont classés dans la population, et leur adaptation n’est plus fonction que du rang qu’ils occupent). On peut également envisager, lors de l’exécution de l’opérateur de croisement, de conserver parmi les deux parents et les deux enfants, les deux individus qui ont la meilleure adaptation. On effectue alors un tournoi entre les parents et les enfants à l’issue du croisement. Le danger de ce principe est la convergence prématurée de la population vers un minimum local.
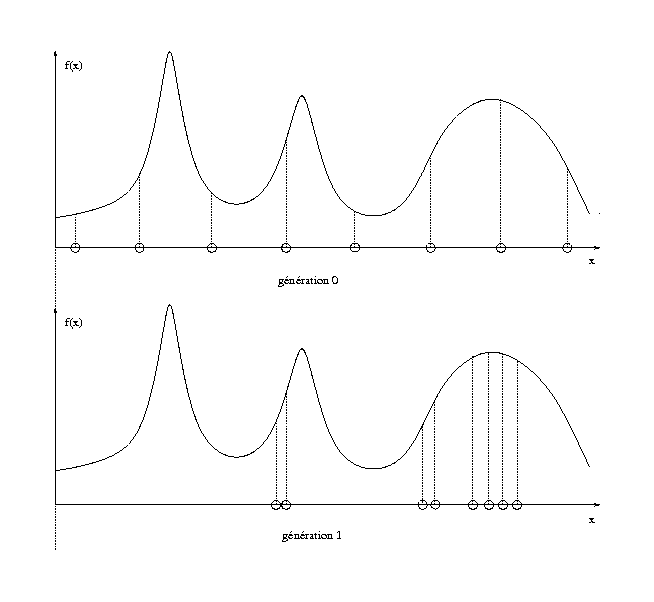
Un des problèmes souvent rencontrés dans l’utilisation des algorithmes génétiques est la disparition trop rapide de certains éléments de la population en raison de la sur-domination d’un individu particulier, dont les représentants finissent par constituer la majeure partie de la population. Considérons l’exemple de la figure 1.7 : à la génération 0, la population de 8 individus est uniformément répartie sur l’intervalle de recherche. Malheureusement, les meilleurs individus ayant une plus forte probabilité de reproduction que les mauvais, on peut se retrouver, à la génération 1 avec une population répartie autour de minima locaux. Le but du sharing est de contrecarrer ce phénomène.
De plus, un des avantages des algorithmes génétiques par rapport aux algorithmes de type recuit simulé est de présenter plusieurs solutions viables, et non une seule, au terme d’une exécution du programme. Il faut donc éviter l’élimination de tous les individus au bénéfice d’un seul, même s’il est mieux adapté.
La technique dite de sharing (que l’on pourrait traduire par partage ou répartition en français) tente de faire disparaître ce problème. Le principe est simple : la fitness de chaque individu est modifiée en fonction du nombre d’individus qui l’entoure dans l’espace du problème ; plus ce nombre est grand, et plus on diminue la fitness. La formule classiquement utilisée est :
| fs(xi)= |
|
fs(xi) est la valeur modifiée de la fitness de l’élément xi après l’application de l’opération de sharing ; f(xi) est la fitness << traditionnelle >> ; d(xi,xj) est la distance séparant les éléments xi et xj ; s est une fonction << astucieusement >> choisie. Cette fonction a souvent la forme triangulaire représentée sur la figure 1.8.
On voit que tout élément situé à une distance supérieure à s0 ne modifie en rien la fitness, mais qu’en revanche, plus il y a d’éléments proches et plus la fitness se trouve diminuée. Cette technique permet ainsi de pénaliser les zones trop peuplées, contenant trop de représentants d’une même solution, ou d’une solution proche ; elle favorise en revanche les représentants uniques ou peu nombreux d’une solution.
Le sharing nécessite la définition d’une distance sur l’espace de recherche et pénalise le temps d’exécution de l’algorithme car si la population est de taille n, le temps de calcul des distances entre les paires d’individus est proportionnel à n2. Il existe des méthodes approchées [YG93] ou permettant de ramener le temps de calcul proportionnel à n log(n).
L’utilisation du sharing sur l’exemple de résolution de conflit permet d’une part d’améliorer la solution obtenue (figure 1.9) et d’autre part d’obtenir plusieurs solutions différentes.
Les algorithmes génétiques sont très souvent utilisés pour des problèmes de grande taille où l’on cherche à minimiser un critère additif, somme de critères faisant chacun intervenir une partie des variables. Ce type de fonction n’est pas marginal, beaucoup de problèmes pratiques faisant intervenir un grand nombre de variables sont partiellement séparables. Nous verrons à la fin de cette section que notre problème de résolution de conflits aériens en fait partie.
L’idéal, pour résoudre un problème à plusieurs variables est de pouvoir séparer le problème en sous-problèmes indépendants. La solution globale est reconstituée à partir des solutions de chacun des sous-problèmes. Considérons l’exemple de la fonction définie sur ℜ n (x1,..,xn)→ ∑1n xi2. Cette fonction est complètement séparable, elle est la somme de n critères positifs xi→ xi2 que l’on peut minimiser indépendamment pour trouver la solution globale. Supposons que l’on n’ait pas profité de cette propriété et considérons l’opérateur de croisement simple d’un AG qui, à partir de deux parents (x1,..,xn) et (y1,..,yn) génère les enfants (z1,..,zn) et (z1,..,zn) avec (zi,zi)=(xi,yi) ou (yi,xi) . Or le meilleur enfant que l’on peut produire est obtenu en choisissant pour zi la variable xi ou yi qui minimise le critère t→ t2.
On peut ainsi largement améliorer la convergence d’un algorithme génétique en utilisant un opérateur de croisement qui tient compte de la séparabilité du critère à optimiser pour fabriquer les enfants à partir de 2 parents.
Par la suite, on appellera fonction partiellement séparable toute fonction positive F de n variables x1, x2, .., xn pouvant s’écrire comme la somme de m fonctions positives Fi, chacune ne faisant intervenir que ni variables (ni<n).
| F(x1,x2,..,xn)= |
| Fi(xj1,xj2,..,xjni) |
On définit l’adaptation ou fitness locale Gk(x1,x2, ..,xn) pour la variable xk comme suit :
| Gk(x1,x2,..,xn)= |
|
|
où Sk est l’ensemble des i tels que xk est une variable de Fi et ni le nombre de variables de Fi.
Intuitivement, l’adaptation locale d’une variable isole sa contribution à l’adaptation globale. On a de plus la propriété suivante :
| Gk(x1,x2,..,xn)=F(x1,x2,..,xn) |
L’opérateur de croisement fabrique un enfant z1,..,zn à partir de deux parents comme suit :
Le paramètre Δ rend plus où moins aléatoire la construction de l’enfant.
Ce principe s’applique très bien au problème de résolution de conflits : en effet, on cherche d’abord à minimiser C somme de critères faisant intervenir des paires d’avions, puis on cherche à minimiser la somme des allongements, critères ne faisant intervenir qu’un avion chacun. Le croisement adapté permet, pour le problème de résolution de conflit, de résoudre des configurations atteignant une vingtaine d’avions, comme le montre la figure 1.10.
Les algorithmes génétiques peuvent optimiser n’importe quel type de structure de données dès lors que l’on est capable de définir une fonction d’évaluation et des opérateurs de recombinaison. On peut ainsi, comme le décrit l’exemple 1.5.7 optimiser les poids d’un réseau de neurone, voire même sa structure. Pour un problème d’optimisation opérant sur des vecteurs réels, on peut définir comme élément de population non plus le vecteur lui-même, mais un simplexe de vecteurs (une méthode locale permettant d’optimiser le simplexe à chaque génération [DA99]). On peut également envisager des éléments de population diploïdes.
Il était inévitable que l’on tente d’utiliser pour les algorithmes génétiques des chromosomes diploïdes en lieu et place des chromosomes haploïdes utilisés jusqu’ici : les organismes vivants les plus évolués ont des chromosomes diploïdes12. Dans ce cas, le décodage du génotype au phénotype se fait en introduisant la notion de dominance d’un gène sur un autre, comme en génétique classique (voir [Gol89c]).
Les diverses expériences faites sur la diploïdie tendent à montrer que cette technique n’est efficace que lorsque la fonction fitness varie en fonction du nombre de générations de l’algorithme génétique i.e., f est de la forme f(C,n) (où n est le numéro de génération de l’algorithme génétique), au lieu d’être simplement de la forme f(C). Ceci semble être en accord avec la génétique qui pense qu’un des facteurs positifs de la diploïdie est sa capacité à mémoriser (par l’intermédiaire de gènes récessifs) des caractéristiques qui ont pu se révéler utiles dans le passé, ne le sont plus aujourd’hui, et donc doivent disparaître du phénotype ; cependant, si une évolution de l’environnement venait à rendre ces caractéristiques utiles à nouveau, les gènes récessifs les codant étant disponibles, elles réapparaîtraient sur le champ.
La diploïdie est, pour l’instant, peu utilisée pour la programmation des algorithmes génétiques.
Nous allons présenter dans cette section une intéressante combinaison reliant les algorithmes génétiques aux réseaux de neurones.
Considérons le problème suivant : nous disposons dans un espace clos de six mobiles. Chacun de ses mobiles à une origine et une destination. Le but de chacun de ces engins est de rejoindre sa destination en suivant le plus court chemin. Cependant, ils ne doivent à aucun moment se trouver à une distance d’un autre mobile inférieure à une norme de séparation D, et ce durant l’intégralité de leur déplacement ; il s’agit donc de construire une trajectoire optimale au sens de la distance tout en respectant des contraintes de séparation.
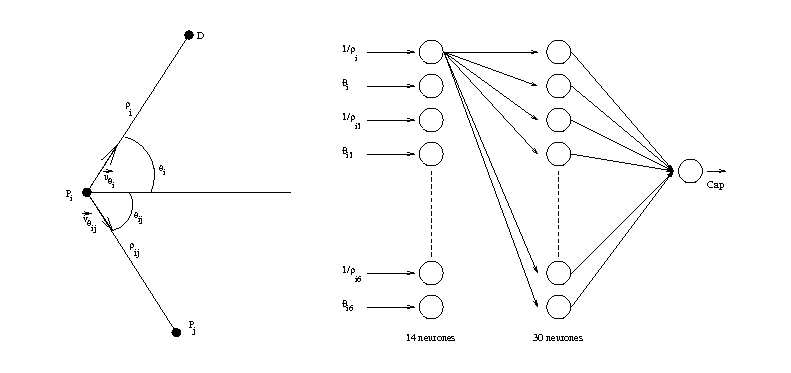
Pour chaque mobile i, les paramètres en entrée sont les distances ρij aux autres mobiles, les gisements de ces mobiles θij ainsi que la distance ρi et le gisement θi de sa destination. En sortie, le programme doit fournir un cap à suivre αi pour le pas de temps suivant (on suppose que les mobiles avancent par pas de temps discret, à une vitesse unique et fixée).
On peut donc construire un réseau de neurones qui aurait pour entrées les différents paramètres et fournirait en sortie le cap du mobile (voir figure 1.11).
Le problème est de réaliser l’apprentissage du réseau. On ne dispose en effet d’aucune base d’exemple permettant de faire de l’apprentissage supervisé, car il n’existe pas de solution mathématique générale à ce problème.
On peut alors utiliser la technique des algorithmes génétiques pour calculer les poids du réseau de neurones. On commence par générer un ensemble de poids pour notre réseau (un chromosome contient en fait la matrice des poids). Puis, on fait se déplacer nos mobiles, sur un exemple ou un ensemble d’exemple, avec les poids de chacun des chromosomes. La fitness de chaque chromosome correspond à la qualité de la trajectoire générée. Le plus court chemin est bien sur l’optimal, mais il faut éviter les autres mobiles. Toute collision pénalise la fitness, soit en l’annulant, soit en la divisant par un coefficient donné.
Les opérateurs de croisement sont du type barycentrique. On tire aléatoirement deux coefficients dans deux réseaux et on réalise une combinaison linéaire pour obtenir les nouveaux coefficients. La mutation consiste à ajouter un bruit blanc sur un coefficient tiré au hasard.
On peut ainsi construire un réseau qui parviendra à réaliser les deux objectifs fixés : de bonnes trajectoires en terme de distance et un évitement des autres mobiles13.
La combinaison des algorithmes génétiques et des réseaux de neurones ouvre une voie particulièrement intéressante pour les problèmes où l’on ne dispose pas de base de données d’exemples permettant de faire de l’apprentissage supervisé.
Il est rare, pour un problème réel, que l’on n’ait qu’un seul critère à optimiser ; bien souvent, on se contente de rechercher un compromis entre plusieurs critères (par exemple une combinaison linéaire entre les critères). Or les algorithmes génétiques font évoluer une population d’individus, et on peut être tenté de faire converger cette population non plus vers un point optimum, mais vers la surface de Pareto des différents critères.
La notion de dominance au sens de Pareto se définit comme suit : on dit que le point A domine le point B si, ∀ i, fi(A)≥ fi(B) (avec au moins une inégalité stricte), où les fi représentent les critères à maximiser. L’ensemble des points qui ne sont dominés par aucun autre point forme la surface de Pareto. Tout point de la surface de Pareto est “optimal”, dans la mesure où on ne peut améliorer la valeur d’un critère pour ce point sans diminuer la valeur d’au moins un autre critère.
On peut construire un algorithme génétique permettant de trouver la surface de Pareto d’un problème multi-critères.
La technique employée dérive directement des travaux de Jeffrey Horn et Nicholas Nafpliotis ([HN93]). Au lieu de travailler sur une fonction d’évaluation scalaire, on a désormais une fonction d’évaluation vectorielle (chaque critère est une variable du vecteur). En conséquence, le principal changement induit concerne le processus de sélection14.
On introduit une variante de la notion de dominance que l’on définit ainsi : l’élément Ei domine Ej si le nombre des valeurs contenues dans son vecteur d’adaptation qui sont supérieures aux valeurs correspondantes dans Ej dépasse un certain seuil. La technique proposée pour effectuer la sélection est alors simple : on tire deux individus au hasard ainsi qu’une sous-population15 à laquelle ils n’appartiennent pas et qui va servir à les comparer.
Trois cas se présentent alors :
Le sharing va conduire à sélectionner celui des deux individus qui a le moins de voisins proches, autrement dit, on élimine celui qui se situe dans une zone d’agrégation pour conserver celui qui est dans une région moins dense.
Le terme voisin proche n’a pas de signification précise mais il est possible de définir un voisinage (aussi appelé niche) correct en se servant de la distance de Holder :
| dH(Ei,Ej)= |
|
|
fik désignant la k-ième composante du vecteur adaptation de l’élément i. Le paramètre p permet de faire varier la forme et la taille du voisinage. A l’intérieur des voisinages ainsi définis dans l’espace objectif, il suffit de compter le nombre d’individus pour favoriser les zones les moins denses, et de cette façon, maintenir la diversité de la population.
Pour certains problèmes d’optimisation, la fonction fitness résulte d’une évaluation sur un certain nombre de cas tests. Or bien souvent, il est difficile de choisir une base de cas tests représentative de tous les cas possibles. Imaginons par exemple que l’on veuille approcher la courbe que nous fournit une boîte noire par un polynôme de degré n et que l’on veuille déterminer les coefficients du polynôme en utilisant un algorithme génétique. On peut sélectionner un certain nombre de points de la courbe qui permettront, pour un polynôme donné de mesurer la qualité du polynôme retenu. Rien ne garantit toutefois qu’une autre sélection de points ne donne pas une évaluation totalement différente.
Le principe de la coévolution est de faire évoluer deux populations différentes en parallèle. Dans le cas présent, la première population est une population d’individus codant les coefficients d’un polynôme, la deuxième population est une population de points de la courbe servant à tester la première population.
Il n’existe pas d’algorithme de coévolution canonique. Différents travaux ayant utilisé le principe de coévolution permettent de donner une image plus concrète de ce que peut être la coévolution :
Dans un article de 1992, Co-evolving Parasites Improve Simulated Evolution as an Optimization Procedure [Hil92], Hillis présente une méthode dont le but est de générer des réseaux de tri, parvenant à trier un nombre n donné de nombres au moyen du plus petit nombre possible de comparaisons successives. Les éléments de la première population sont des réseaux de tri, qui évoluent au moyen d’un algorithme particulier. Nous développerons les caractéristiques de cet algorithme qui nous semblent jouer un rôle important dans l’utilisation par Hillis de la coévolution. Les parasites sont des ensembles de cas tests, c’est-à-dire des séquences d’entiers (de 0 et de 1 puisque un réseau de tri qui trie correctement toute séquence de 0 et de 1 de taille n trie correctement toute séquences de nombres de taille n). Les réseaux ont une adaptation d’autant plus grande qu’ils parviennent à ordonner correctement une plus grande proportion de ces cas tests La coévolution consiste dans ce cadre a faire évoluer, en même temps que la population de réseaux, une population de parasites.
Hillis obtient grâce à la coévolution de meilleurs résultats que ceux qu’il avait obtenus au moyen de l’évolution simple des réseaux, tant en termes d’efficacité des réseaux pour un plus petit nombre de comparaisons, qu’en terme de rapidité d’obtention de ces réseaux. Il voit à ce résultats deux explications. D’abord, les réseaux imparfaits (qui ne trient pas correctement toutes les séquences de n entiers), qui pourraient constituer des optima locaux sont la proie de parasites adaptés à leurs faiblesses qui se développent dans leur voisinage, et ainsi ne sont pas viables (on voit ici le rôle de la localisation des éléments des deux populations sur une grille). De plus, les cas tests les plus difficiles dominent rapidement dans la population des parasites, ce qui évite des évaluations inutiles de réseaux sur des cas tests trop faciles.
Les stratégies d’évolution sont en fait généralement désignées, même dans les milieux francophones, par leur nom anglais de Evolution Strategies, ou par le sigle ES. Elles ont été introduites par Schwefel, à Berlin en 1965. à l’origine, un individu unique produisait, au moyen d’une mutation, un descendant unique, une sélection s’opérait entre eux deux.
Dès leurs origines, une grande originalité des stratégies d’évolution par rapport aux algorithmes génétiques a résidé dans leur utilisation du codage réel (voir section 1.5.1) et de la mutation gaussienne (section 1.5.1).
Elles ont été étudiées et développées par Rechenberg, à qui on doit la règle dite du un cinquième : le taux de mutation est modifié au cours du déroulement de l’algorithme de manière à atteindre un taux de succès des mutation de un cinquième, c’est-à-dire que dans un cas sur cinq en moyenne, le produit d’une mutation (le fils) a une meilleure fitness que l’élément à partir duquel il a été obtenu (le père). Des techniques similaires ont pu ensuite être reprises dans le cadre d’autres algorithmes évolutionnaires, de même que la self adaptation, introduite par Schwefel (voir section 1.5.1).
Dans les années 70 ont été développées 2 versions des stratégies d’évolution, les (µ+λ)-ES et les (µ,λ)-ES, dues à Schwefel, dont on trouvera une description dans [BS93]. Dans les deux cas, µ individus donnent, au moyen de mutations (et éventuellement d’opérateurs de recombinaisons, tels que le croisement décrit dans la section 1.5.1), λ nouveaux individus. Dans le cadre des (µ,λ)-ES, λ > µ, et la sélection retient µ individus sur les λ qui ont été créés. Elle retient, dans le cadre des (µ+λ)-ES, µ individus sur l’ensemble des µ parents et des λ enfants. La sélection est déterministe : les meilleurs λ individus (en termes de fitness) sont conservés, les λ moins bons sont éliminés.
Les stratégies d’évolution, en plus de former un groupe d’algorithmes très intéressant en eux mêmes, ont eu une influence positive sur développement des algorithmes génétiques, notamment par la mise au point de nouveaux opérateurs de mutation, et par l’expérience du codage réel qu’elles ont permis d’acquérir.
Le principe de la programmation évolutionnaire, ou Evolutionnary Programming a d’abord été présenté par L. J. Fogel [FOW66] en 1966.
Ces algorithmes ont été grandement développés (notamment par le passage au codage réel) et popularisés par D. B. Fogel [FFAF92].
Leurs principales caractéristiques, par rapport aux autres algorithmes évolutionnaires, étaient, à l’origine, le fait qu’ils n’utilisaient pas de croisement, et surtout l’utilisation d’un mode de sélection originale, la sélection par tournoi.
Nous donnons ici une description simple d’un algorithme de programmation évolutionnaire (cette famille d’algorithmes s’est rapidement enrichie et diversifiée). µ individus en donnent µ par mutation. Si le codage utilisé est un codage réel, cette mutation est gaussienne, et son écart-type peut, au moment de la mutation d’un individu, être calculé à partir de la fitness de cet individu.
Notons P(n) la population des µ individus présent à la génération n, et P′(n) les µ individus obtenus à partir d’eux par mutation. On va d’abord attribuer une note à chacun des 2 µ individus de P(n) ∪ P′(n). Pour chaque individu e de P(n) ∪ P′(n), on tire aléatoirement q individus dans P(n) ∪ P′(n). La note de e est alors égale au nombre d’individus (parmi ces q) dont la fitness est inférieure à celle de e.
La sélection retient pour former la nouvelle population P(n+1), les µ éléments ayant obtenu les meilleures notes.
Les performances de représentants de ces deux familles d’algorithmes (ES et EP), et d’un algorithme génétique (utilisant un codage binaire) sont comparées sur des problèmes d’optimisation de fonctions test de ℜ n dans ℜ , dans [BS93].
La programmation génétique est une technique récente qui cherche à appliquer les techniques génétiques sur des ensembles de programmes et non sur des données. Nous ne décrirons que très brièvement la programmation génétique, le lecteur intéressé devra se reporter à [Koz92] : une présentation exhaustive, accompagnée de nombreux exemples, des techniques de programmation génétique.
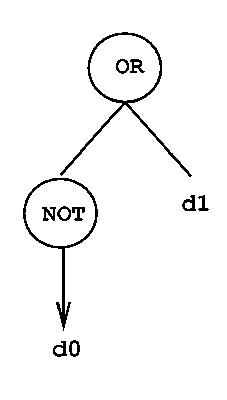
Prenons un exemple simple : supposons que nous souhaitions construire une fonction LISP à partir des fonctions logiques classiques (AND, OR, NOT) qui implante l’implication. Il est clair que dans ce cas, la fonction s’écrit simplement :
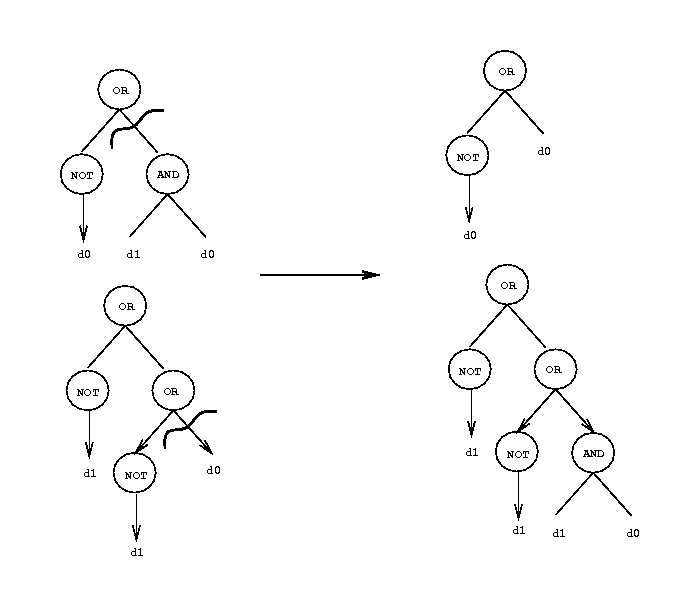
La programmation génétique permet de générer automatiquement une telle fonction à partir de la table de vérité de l’implication. Le fonctionnement est le suivant :
Un des problèmes principaux de la programmation génétique est le choix des opérateurs à introduire dans la population initiale. Dans notre cas, le choix de NOT, AND et OR était relativement simple à faire. Dans bien d’autres cas, il n’en est pas de même, et le choix initial des opérateurs conditionne bien souvent la réussite ou l’échec du mécanisme de programmation génétique. Comme dans bien d’autres cas, le savoir-faire de la personne qui met la technique en œuvre conditionne le résultat bien plus sûrement que la technique elle-même.
Au même titre que les réseaux de neurones, bien que dans une optique différente, les algorithmes génétiques représentent une approche originale et intéressante. Là aussi, il faut se garder de se laisser prendre aux phénomènes de mode et songer à comparer les algorithmes génétiques à des méthodes plus classiques d’optimisation (recuit simulé, branch and bound, etc.) On peut considérer qu’ils sont passés du stade de la recherche pure à celui de la recherche appliquée.
Le téléchargement ou la reproduction des documents et photographies
présents sur ce site sont autorisés à condition que leur origine soit
explicitement mentionnée et que leur utilisation
se limite à des fins non commerciales, notamment de recherche,
d'éducation et d'enseignement.
Tous droits réservés.
Dernière modification: 09:48, 22/02/2024