La perception des couleurs chez l’être humain |
Ce travail a été effectué par mon fils et trois de ses camarades (Antoine Brunet, Thomas Dalla Verde et Aubin Maria) dans le cadre d’un TPE de première. Mon travail dans cette affaire a été de leur fournir du matériau sur lequel travailler, les guider et les aider à la mise en forme finale. Il ne faut évidemment pas en attendre plus que ce qu’il est, et il existe certainement des erreurs ou des omissions. Cependant, il s’agit me semble-t-il d’une introduction intéressante au sujet.
La vision de la couleur est une activité complexe, et est, de plus une activité totalement dépendante de l’observateur. Comme nous allons le voir, il n’existe à proprement parlé dans la nature qu’un nombre “réduit” de couleurs existant objectivement: les couleurs monochromatiques. Toutes les autres couleurs sont des créations du système nerveux et du cerveau. Les problèmes de vision de la couleur peuvent donc avoir leur origine soit dans des dysfonctionnements au niveau de l’œil, soit au niveau cérébral.
Les mécanismes de la vision des couleurs sont relativement bien connus aujourd’hui en ce qui concerne la physiologie de l’œil et des récepteurs spécifiques qu’il contient. En revanche les mécanismes qui sont mis en oeuvre dans le cerveau sont beaucoup moins bien connus et les théories scientifiques dans ce domaine évoluent très vite.
Dans le chapitre 2, nous allons introduire rapidement ce qu’est la lumière, quelles sont ses propriétés relatives à la couleur et à sa vision et Nous présenterons ensuite l’évolution historique des théories relatives à la vision de la couleur. Dans le chapitre 3, nous détaillerons l’anatomie de l’œil. Puis dans le chapitre 4, nous développerons les principes de la vision dite trichromique, ses fondements, ainsi que les validations expérimentales du modèle trichromique. Dans le chapitre 5, nous poserons les bases génétiques et biologiques qui sous-tendent le fonctionnement de l’œil humain, nous expliquerons comment ces résultats sont en accord avec le modèle trichromique et comment ils expliquent la majorité des différences dans la perception de la couleur. Enfin, dans le chapitre 6 nous montrerons comment le traitement cérébral de l’information est central dans l’appréhension de la couleur, et comment il permet également d’expliquer des différences d’appréciation des couleurs que l’on ne peut pas comprendre avec le modèle trichromique. Nous terminerons en discutant des problèmes pratiques liés aux défauts de la vision de la couleur dans le chapitre 7.
Ce chapitre est un résumé rapide du problème. Une version plus longue se trouve ici.
Deux théories se sont longtemps opposées concernant la nature de la lumière. La première, la théorie corpusculaire, défendue par Newton, soutenait que la lumière était composée d’un ensemble de particules. La seconde (la théorie ondulatoire), défendue par Huyghens et Fresnel, présentait la lumière comme une onde qui servait de vecteur aux champs électriques et magnétiques.
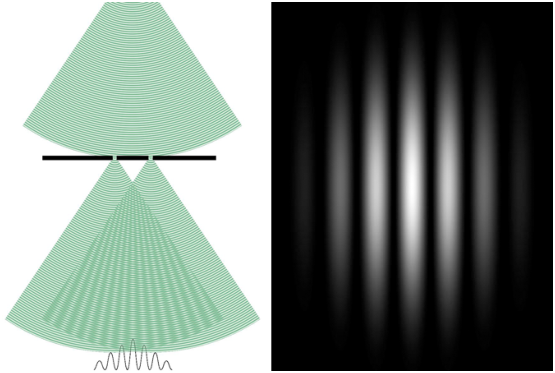
Au début du XIXème siècle, la théorie ondulatoire semble s’imposer: Thomas Young réalise l’expérience des fentes doubles (voir figure 2.1), qui montre que la lumière interagit comme une onde en créant des figures d’interférence. Quelques années plus tard, James Clerk Maxwell unifie le magnétisme, l’électricité et les différentes ondes, montrant aussi que la lumière est une radiation électromagnétique, qui est donc caractérisée par sa longueur d’onde.
Pourtant, au début du XXème siècle, un certain nombre de scientifiques, donc Einstein, Planck ou de Broglie montrent que l’interprétation purement ondulatoire de la lumière est insuffisante: ils posent les bases de la mécanique quantique qui dit que toute particule exhibe les deux types de comportement, à la fois corpusculaire et ondulatoire.
Montrer de façon expérimentale les deux aspects corpusculaires et ondulatoires de la lumière est difficile, et il a fallu plusieurs années avant qu’une expérience montre de façon très “visuelle” les effets quantiques de la lumière.
En 1989, une expérience réalisé par le professeur Tanamura permettra de parfaitement visualiser ce double aspect de la lumière. En bombardant individuellement avec des photons une plaque, elle permet de montrer l’apparition lente des figures d’interférence de Young (voir figure 2.2).
La lumière est donc une onde dont le vecteur est une particule élémentaire appelée photon. Les photons sont des particules sans masse qui ont une énergie E=h ν= h c/λ ou h est la constante de Planck, c la vitesse de la lumière, ν la fréquence et λ la longueur d’onde.
Les photons sont libérés par les atomes lorsqu’un électron qui a été excité retombe dans son état d’énergie initial. La longueur d’onde du photon émis dépend de l’écart d’énergie entre l’état de départ et l’état final (ou fondamental) de l’électron suivant la formule λ = Ed − Ef/h c. La théorie quantique ayant établi que les électrons ne peuvent occuper que des états d’énergie discrets dans un atome, les longueurs d’onde (et donc la lumière) émises par un atome ne sont pas continues, mais correspondent à des bandes, appelées bandes spectrales, qui permettent d’ailleurs d’identifier ces atomes, par exemple en astronomie.
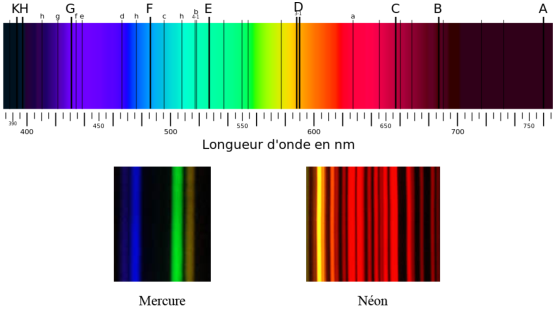
Ce phénomène avait jusque là été mal compris; Newton le premier avait découvert que la lumière blanche se décomposait avec un prisme en un ensemble de couleurs qui restaient les mêmes quelle que soit l’orientation du prisme ou son matériau. Mais la première observation d’une raie spectrale est faite pas Thomas Melvill en 1752 (le jaune du sodium). Ces observations seront complétées au fil des années par William Wollaston, Joseph Von Fraunhofer, Gustav Kirchhoff et Anders Angstrom. On peut voir sur la figure 2.3 les spectres respectifs du mercure et du néon, ainsi qu’une décomposition spectrale complète du spectre visible telle que faite par un prisme.
Le mercure a un spectre relativement simple avec seulement cinq bandes de longueur d’onde 404,7 nm, 435,8 nm, 546,1 nm, 577 nm et 579,1 nm. Celui du néon est beaucoup plus complexe, avec de très nombreuses bandes toutes situées dans le jaune et le rouge.
Nous avons parlé jusqu’ici de sources émissives. En ce qui concerne les matériaux qui se contentent de refléter la lumière (ou que la lumière traverse), le mécanisme est presque identique: ces matériaux vont absorber une partie des photons qu’ils reçoivent, mais ils ne pourront absorber que les photons dont l’énergie correspond à un écart de niveaux d’énergie possible dans cette molécule. Par exemple, le dioxyde de carbone CO2 à un écart minimal d’énergie égal à environ 8eV (c’est l’écart entre le dernier niveau d’énergie occupé par un électron dans la molécule au repos et le premier niveau libre supérieur d’énergie). Cela correspond à un photon situé dans l’ultraviolet: les molécules de dioxyde de carbone absorbent l’ultraviolet, mais laissent passer toutes les radiations du spectre visible à l’œil humain: elles nous sont donc invisibles. Il en va de même pour de nombreuses molécules “simples” comme le dioxygène ou l’azote. Il existe en revanche des molécules organiques plus complexes qui contiennent une alternance de liaisons carbone simples et de liaisons carbone doubles, ou liaisons conjuguées. Ces molécules ont des niveaux d’absorptions situés dans le spectre visible et elles sont la plupart du temps la cause de la pigmentation des objets organiques; on les appelle des chromophores, et nous en reparlerons dans la section consacrée à la physiologie de l’œil1.
Dans l’absolu, les couleurs qui composent le spectre visible représenté sur la figure 2.3 sont les seules couleurs “réelles”. Chacune d’entre elle est une couleur dite monochromatique, c’est à dire qu’elle est entièrement définie par une seule longueur d’onde. Ces couleurs sont les seules à avoir une existence “objective”. Pourtant, dans la vie de tous les jours, nous “voyons” bien d’autres couleurs, qui n’existent pas “vraiment”. Il s’agit là d’une propriété “subjective”, qui est lié à notre système nerveux et à sa façon d’appréhender les radiations lumineuses.
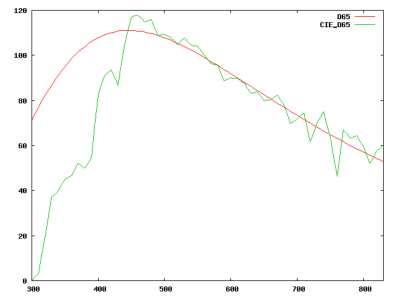
Comme l’avait démontre Newton, une radiation lumineuse est entièrement déterminée par son spectre de puissance. On peut voir sur la figure 2.4 la représentation du spectre de puissance de la lumière “blanche” du soleil reçue sur terre après filtrage par l’atmosphère terrestre, et l’émission d’un corps noir à une température de 6500K. Pour chaque longueur d’onde, on représente la puissance émise pour cette longueur d’onde. C’est le type d’analyse qu’effectue un spectromètre, et c’est la seule définition correcte d’un rayonnement lumineux. Nous allons voir que l’œil humain est bien incapable de fonctionner ainsi.
L’homme a été très tôt conscient de plusieurs phénomènes curieux concernant sa vision des couleurs.
Les peintres (et la peinture est un des arts les plus anciens de l’humanité) savaient bien avant qu’il existe une théorie de la couleur qu’il est possible d’obtenir certaines teintes en mélangeant d’autres couleurs qui n’ont en apparence rien à voir avec elles. Ils réalisaient sans le savoir les premières synthèses soustractives.
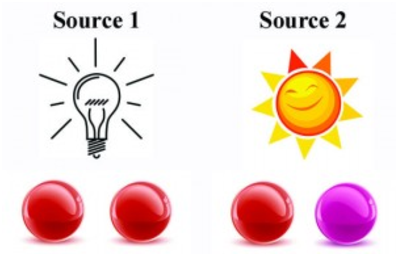
Un des paradoxes les plus anciens connus est le métamérisme qui peut se manifester de diverses façons. La première version du métamérisme est celui qui est représenté sur la figure 2.5: il s’agit de deux objets (différents) qui semblent avoir exactement la même couleur sous un certain éclairage, mais qui ont des couleurs différentes avec un éclairage différent.
La seconde version, tout aussi surprenante, est qu’un même objet peut avoir des couleurs différentes suivant des éclairages qui semblent pourtant être de la même “couleur”. Ces phénomènes ne peuvent s’expliquer en termes de longueur d’onde: un spectromètre ne serait d’ailleurs pas trompé par les exemples ci-dessus. Dans le premier cas, il distinguerait que les deux objets émettent bien des rayonnements différents quel que soit l’éclairage, et dans le second cas il distinguerait clairement les deux éclairages, qui ne sont semblables que pour un œil “humain”.
Le premier a avoir commencé à théoriser ce phénomène est là encore Newton qui dès 1666 se rendit compte que certaines couleurs comme le magenta ou le pourpre ne se trouvaient pas dans le spectre de décomposition de la lumière blanche, et ne pouvaient donc s’obtenir que par un mélange émissif de couleurs monochromatiques; il développa alors la première théorie de la synthèse additive.
L’idée que le système de vision humain est trichromique remonte presque simultanément à James Clerk Maxwell (1855) et à Herman Gunther Grassman (1853). Ces lois, connus sous le nom de principe de composition linéaire ou de lois de Grassman ont posé les principes de la vision humaine des couleurs. Elles sont au nombre de trois:
C’est Herman Von Helmholtz qui développera complètement le modèle trichromique à partir de 1859. Les lois de Grassman et leur vérification expérimentale permettront le développement de tous les systèmes de reproduction additif de la photographie couleur (1861) à la télévision et la photographie numérique, en passant par le cinéma.
Pourtant, dès 1860, Ewald Herring note que ce modèle est imparfait. Herring constate expérimentalement que l’homme est incapable de se représenter certaines couleurs. Par exemple, alors que l’on peut parfaitement imaginer un bleu-vert (c’est d’ailleurs un nom de couleur régulièrement employé), il est impossible de se représenter un rouge-vert (ou même un jaune-bleu). Or le rouge et le vert sont très proches au sens physique du terme (longueur d’onde), au moins autant que le bleu et le vert. Rien dans la théorie trichromique ne permet d’expliquer ce phénomène. D’autre part, si l’on regarde pendant une longue période un carré jaune et que l’on regarde immédiatement après une feuille blanche, on voit apparaître sur celle-ci un carré bleu (voir figure 2.6), ce que la théorie trichromique n’explique pas non plus.

Figure 2.6: En regardant fixement la partie gauche de la feuille, puis rapidement la partie droite, on constate que l’étoile de droite est bleutée au lieu d’être blanche (extrait de [Con09])
Il y a pour Herring des couleurs qui sont en opposition (jaune/bleu, vert/rouge), et le nombre de primaires doit être plus élevé que trois.
On a longtemps cru les deux théories incompatibles, et au milieu du XXème siècles les découvertes sur la physiologie de l’œil qui confirmaient la théorie trichromique ont plongé la théorie de Herring dans l’oubli. Pourtant, quelques années plus tard, les avancées de la biologie ont montré que la théorie de Herring était partiellement valide, et qu’elle expliquait la première étape de traitement de l’information visuelle par le système nerveux.
Enfin, il y a d’autres illusions visuelles tout autant inexplicables par la théorie classique, et même par la théorie de Herring. Le cerveau a ainsi une capacité exceptionnelle à gommer les différences d’éclairage. Par exemple, que l’on regarde une pomme au soleil ou sous un éclairage fluorescent, elle semblera de la même couleur alors qu’un spectromètre (ou un appareil photo!) mesurerait des valeurs extrêmement différentes. Ce mécanisme, appelé constance de la couleur (“color constancy” en anglais), est fortement implanté dans le traitement cérébral de l’information. Il est d’ailleurs possible de le tromper en construisant des scènes où ces capacités inconscientes d’interprétation sont prises en défaut, à tel point qu’il est impossible de s’en défaire même lorsqu’on connaît le “truc”, comme sur la figure 2.7: les trois facettes marquées d’une étoile sont techniquement exactement de la même couleur: grises. On peut s’en convaincre avec un logiciel de traitement d’image en mesurant les composantes RGB des trois facettes. Pourtant, elle parait jaune dans un environnement bleu, bleue dans un environnement jaune et grise seulement dans un environnement blanc. Il est impossible d’échapper à l’illusion, même en essayant consciemment d’y résister.
La physiologie de l’œil ne commencera à être bien connu que dans les années 50 à partir des travaux de Gunnar Svaetichin. On voit sur la figure 3.1 la structure simplifiée de l’œil. Nous avons eu l’occasion d’effectuer lors d’un TP une dissection de l’œil de veau, et nous inclurons dans ce chapitre un certain nombre de photos réalisées lors de cette dissection.
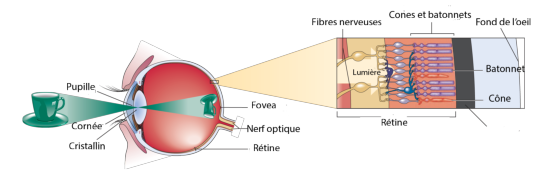
Figure 3.1: Structure de l’œil (à partir de [Gol10])
L’œil est l’organe de la vision, qui permet à un être vivant de capter la lumière pour ensuite l’analyser et interagir avec son environnement. Il permet de distinguer les formes et les couleurs, tout comme les différentes nuances.
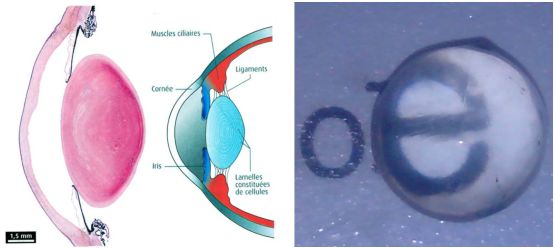
L’œil, ou globe oculaire mesure à peu près 2,5 cm de diamètre pour un poids d’environ 8 grammes. Il est constitué de 3 tuniques qui forme l’enveloppe externe de l’œil de façon analogue à la membrane plasmique d’une cellule. On distingue:
La plus grand partie de l’œil est constituée par l’humeur vitrée qui maintient l’équilibre de l’œil, les différentes parties de l’œil en place. L’œil est constitué de 2 milieux: un milieu transparent , qui laisse passer la lumière et les couleurs dans l’œil et un milieu opaque qui régule la transmission lumineuse.
La pupille, l’iris, la cornée et le cristallin forment un système optique dont le but est de maintenir une image nette sur la rétine, et de réguler également la quantité de lumière qui va l’atteindre.
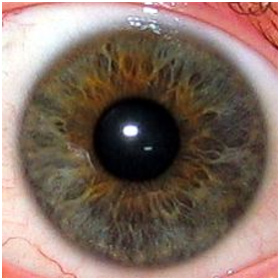
L’iris est la partie colorée de l’œil. C’est un voile très fin formé de lamelles pigmentaires qui donnent leurs couleurs aux yeux. Plus ces pigments contiennent de la mélanine ,qui a pour but de colorer, plus l’œil est foncé. Il règle la dilatation de la pupille en fonction de la luminosité reçue (voir figure 3.2). Pour certains yeux verts, la molécule responsable de la couleur est la lipofuscine. En l’absence de pigments (albinisme), l’œil prend la couleur du sang qui y circule: c’est à dire rouge. Il existe toutes les nuances possibles et imaginables de couleur d’iris, des plus communes aux plus rares. Certains sont même composés d’une multitude de couleurs, et donc distinguer la couleur dominante est parfois très difficile. Il est d’ailleurs parfois impossible de discerner une couleur dominante tant il y a de nuances.
Le cristallin est un des milieux transparents constituant l’œil. Il a une forme de lentille biconvexe. Placé juste derrière la pupille, le cristallin fonctionne comme une loupe très souple directement intégrée dans notre œil. Lorsque l’on fixe un objet au loin, le cristallin est au repos, légèrement bombé : il permet à l’image de se reconstituer sur la rétine, comme sur un écran. Si on fixe un objet de près, l’œil règle la netteté de l’image : le cristallin s’arrondit davantage, pour que les rayons lumineux se rencontrent, à nouveau, sur la rétine. Enfin si le cristallin adopte sa forme normale, l’objet observé se situe à la distance maximale que peut supporter l’œil Nous n’avons pas le temps de nous apercevoir que notre œil a rétabli la netteté de l’image: cette mise au point,s’appelle l’accommodation. L’accommodation du cristallin est rendue possible grâce aux cellules qui le compose: elles ont perdu leur noyau, sont remplies de cristalline (protéine soluble transparente) et sont également très élastiques. L’accommodation est régulée par les muscles ciliaires, situés sur la tunique moyenne.
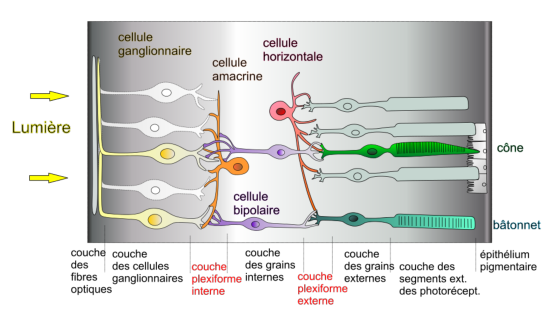
La rétine est le siège principal de la vision. C’est une mince membrane d’environ 0,5 mm d’épaisseur couvrant environ 75% de la face interne du globe oculaire. Elle est principalement constituée de cellules photoréceptrices, appelées cônes et bâtonnets qui analysent la lumière qui arrive après avoir été focalisée et filtrée par la cornée et la pupille. Une autre partie importante de la rétine se consacre au traitement de l’information venue des récepteurs; nous la décrirons plus en détail dans la section 6.3 lorsque nous parlerons du traitement cérébral de l’information lumineuse.
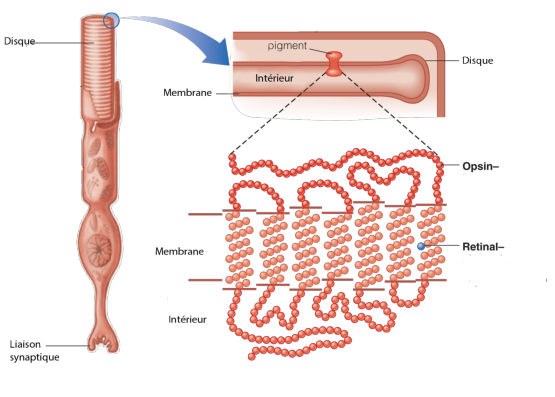
La structure globale des cônes et des bâtonnets est à peu près la même; on voit sur la figure 3.4 la structure précise d’un bâtonnet.
La liaison synaptique s’effectue par le bas vers un neurone bipolaire. Le noyau de la cellule se trouve au dessus de la liaison synaptique. le segment moyen contient les mitochondries et les autres éléments fonctionnels de la cellule (appareil de Golgi,...) Le segment extérieur comporte une série de disques qui contiennent les molécules effectivement sensibles à la lumière. Les cônes ont une structure physique à peu près équivalente: la seule différence visible est leur forme: ils sont un peu plus petits, et en forme de cônes dans leur partie supérieure, d’où leur nom.
L’élément sensible à la lumière dans les bâtonnets est la rhodopsine (dans les cônes il s’agit de la photopsine, que l’on appelle aussi parfois iodopsine). Il n’existe qu’un seul type de rhodopsine, dont la sensibilité spectrale se trouve globalement dans le bleu (autour de 500nm). La rhodopsine est insensible à des longueurs d’onde supérieures à 620nm. La sensibilité spectrale dépend de l’opsine; des opsines différents auront des sensibilités spectrales différentes.
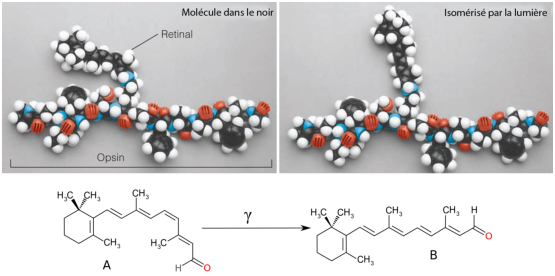
Les opsines sont de très longues protéines qui porte une molécule appelée rétinal (ou retinaldéhyde, ou vitamine A aldéhyde). Il s’agit d’un chromophore, une molécule contenant des liaisons conjuguées, capable d’absorber des électrons dans le spectre visible (nous en avons déjà parlé précédemment). On peut voir sur la figure 3.5 la structure d’une opsine portant une molécule de rétinal.
En l’absence de lumière, le rétinal existe sous forme cis. Lorsqu’il reçoit un photon, il s’isomérise en forme trans. C’est cette réaction qui va entraîner le blocage de l’émission du neuro-transmetteur de la cellule. En effet, chez les vertébrés, et l’homme en particulier, les cônes et les bâtonnets sont des cellules inhibitrices: elles libèrent un neuro-transmetteur quand elles sont au repos, et elles cessent de le transmettre lorsqu’elles sont excitées.
Les bâtonnets sont plus nombreux que les cônes (90 millions contre 5 millions environ). Ils sont beaucoup plus sensibles à la lumière (un bâtonnet peut être excité par un seul photon, alors qu’il en faut plusieurs dizaines pour un cône). Ils disposent de chemins neuronaux différents, qui leur permettent en particulier d’amplifier encore davantage le signal. En particulier les signaux issus des bâtonnets ont un temps d’accumulation de l’ordre de 100ms, bien supérieur à celui des cônes, ce qui permet encore d’améliorer la sensibilité à la lumière. Les bâtonnets ne sont pas impliqués dans la vision de la couleur, ils servent exclusivement à la vision nocturne (“la nuit tous les chats sont gris”). Leur sensibilité spécifique à la lumière est à l’origine du procédé dit de la “nuit américaine” qui consiste à filmer en plein jour avec un filtre bleu pour donner l’impression qu’il fait nuit.
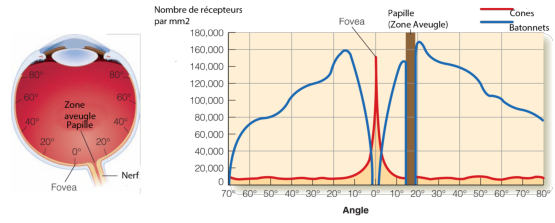
Les cônes se trouvent presque exclusivement dans une zone très petite de l’œil, la fovéa, qui se trouve au centre de la rétine. La densité maximale des cônes dans la fovéa explique que notre vision centrale est particulièrement sensible à la couleur.
On peut voir sur la figure 3.6 la répartition comparée des bâtonnets et des cônes à l’intérieur de la rétine. Contrairement aux bâtonnets, les cônes n’ont pas tous les mêmes opsines ce qui entraîne des différences dans la sensibilité spectrale des cônes.
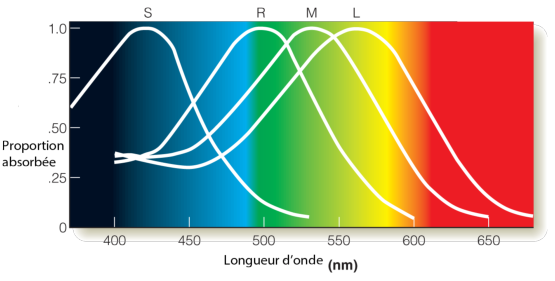
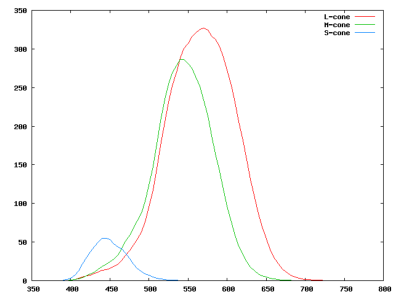
L’œil humain contient principalement trois types de cônes différents, en plus des bâtonnets. Ces cônes sont généralement appelés S (pour Short wavelength, avec une sensibilité spectrale autour de 420–440nm), M (pour Medium wavelength avec une sensibilité spectrale autour de 534–545nm) and L (pour Long wavelength avec une sensibilité spectrale autour de 564–580nm). Nous verrons dans la section suivante quelles ont été les origines génétiques de ces différentes cellules au cours de l’évolution et comment ces origines expliquent en partie leurs propriétés physico-chimiques.
Les cônes L et M ont des sensibilité spectrales relativement proches. Sur la figure 3.7, on peut voir comment l’excitation des quatre types d’opsine de l’œil évolue en fonction de la longueur d’onde du rayonnement (on a également représenté la sensibilité de la rhodopsine (R) en plus des trois types de cônes L, M et S).
La vision humaine est dite trichromique car l’œil humain contient trois types de récepteurs différents. Dans la prochaine section, nous verrons que cela est “presque” équivalent à dire qu’il suffit de trois primaires pour “reconstruire” l’ensemble des couleurs que l’œil humain peut différencier, et nous verrons aussi que cela explique tout un ensemble de propriétés de la vision humaine, même si le simple modèle trichromique est incapable d’en expliquer certaines.
On a longtemps cru que la répartition des différents types de cônes dans la rétine était à peu près régulière et uniforme. On sait maintenant grâce aux progrès de la biologie que ce n’est pas le cas. Il y a beaucoup moins de cônes S que de cônes L et M; la répartition spatiale des cônes dans la rétine n’est pas régulière et on rencontre de façon aléatoire des zones contenant presque exclusivement des cônes L et des zones contenant presque exclusivement des cônes M (voir figure 3.8).
D’autre part, si le ratio de cônes S est approximativement constant d’un individu à l’autre (environ 5% du total), le ratio relatif des cônes L et M peut changer de façon considérable de 80%/20% à 50%/50%. On a montré expérimentalement que ces variations ne changeaient pas la capacité des sujets à distinguer les différentes couleurs. En revanche, rien ne dit (car ce n’est pas un observable objectif) que la sensation de couleur ressentie ne varie pas d’un individu à l’autre: de la même façon qu’il est impossible d’expliquer à un aveugle de naissance ce qu’est la couleur rouge, la sensation de couleur n’est pas partageable entre individus.
Les cônes et les bâtonnets ne sont pas les seules cellules de l’œil à être photosensibles. On sait maintenant depuis une dizaine d’années que l’œil des vertébrés et des invertébrés contient également un type particulier de cellules, appelées cellules ganglionnaires photosensibles, qui sont également capables de générer une réponse à la lumière.
Ces cellules sont très différentes des cônes et des bâtonnets. Elles sont communes aux vertébrés et aux invertébrés. L’opsine qu’elles contiennent (la mélanopsine) fonctionne différemment de la rhodopsine et de la photopsine, et le gène OPN4 qui la code appartient à une autre famille de gènes que celles que nous allons examiner dans le chapitre suivant.
Nous reparlerons un peu du rôle de ces cellules dans le chapitre consacré au traitement de l’information par le cerveau. Elles ne sont apparemment pas impliqués dans le processus de la vision, mais dans celui des rythmes circadiens, même si les scientifiques n’excluent pas complètement qu’elles puissent aussi avoir un rôle pour la vision, comme les cônes et les bâtonnets ont un rôle dans la détermination des rythmes circadiens.
Cela montre une fois de plus la grande complexité de la vision. La figure 3.9 montre le positionnement de ces cellules dans la rétine.
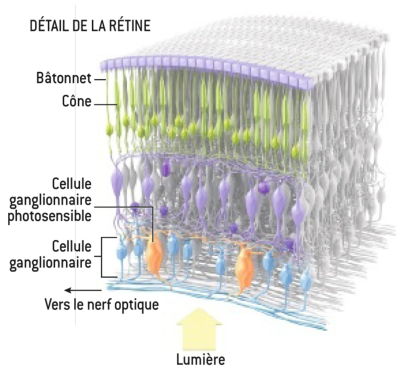
Figure 3.9: Représentation complète des cellules photosensibles de la rétine incluant les cellules ganglionnaires photosensibles (extrait de [Pro12])
Pour plus de détails sur le modèle trichromique, voir ici. Les résultats évoqués ci-dessus sont en total accord avec la théorie trichromique de Maxwell et Van Helmholtz. Elle explique en particulier complètement le métamérisme et les raisons de la reconstruction additive par trois couleurs primaires, que nous allons rapidement expliquer.
Comme nous venons de le voir, l’œil comporte trois types de cônes dont les courbes de sensibilité sont données par la figure 4.1.
Cette figure trace les sensibilités des trois types de cône pour une même quantité d’énergie reçue en fonction de la longueur d’onde. On voit que les cônes bleus sont moins sensibles que les cônes verts et rouges.
Si maintenant nous avons une lumière quelconque, elle peut être entièrement définie par son spectre f(λ) qui définit la puissance émise pour chacune des longueurs d’onde λ. Dans ce cas, la réponse des trois types de cônes (l,m,s) va être:
|
Ces équations permettent de comprendre le métamérisme: en effet, deux couleurs émettant avec des spectres f(x) et g(x) seront vus par l’œil humain comme totalement identiques si les excitations des cônes (l(f),m(f),s(f) et (l(g),m(g),s(g)) sont identiques.
Ces courbes permettent aussi de calculer quelle va être la réponse des trois types de cône à une couleur donnée. Par exemple, pour trois couleurs de longueur d’onde 435, 545 et 700nm, les réponses des cônes vont être:
| L | M | S | |
| λb=435nm | 11.14934 | 14.90023 | 50.30185 |
| λg=545nm | 300.36447 | 286.66280 | 0.17649 |
| λr=700nm | 1.92811 | 0.10506 | 0 |
Si maintenant nous émettons avec ces trois couleurs primaires, et avec des intensités (r,g,b) pour respectivement les composantes rouge (700nm), verte (545 nm) et bleue (435nm), la réponse des cônes (l,m,s) va être:
|
On peut considérer ces équations comme un système de trois équations à trois inconnues, où les inconnues sont les variables (r,g,b) et les données sont les excitations des cônes (l,m,s). En résolvant ce système, on obtient:
|
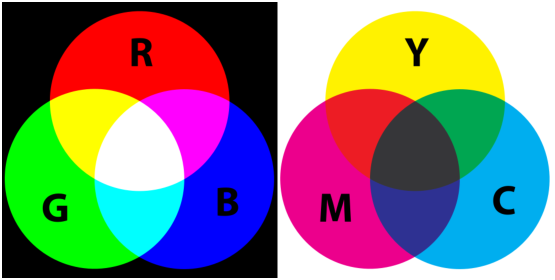
On peut donc reconstruire n’importe quelle excitation des cônes (l,m,s) à partir de trois couleurs (ici monochromatiques) seulement, que l’on appelle en général couleurs primaires. Comme c’est l’excitation des cônes de l’œil qui crée la sensation de couleur, on peut reconstruire pour l’être humain n’importe quelle couleur à partir de trois primaires. C’est sur ce principe que fonctionne tous les systèmes de reconstruction de la couleur dits additifs, c’est à dire des systèmes où les radiations s’additionnent. Un bon exemple est celui des postes de télévision ou des écrans d’ordinateur.

Figure 4.2: Photographie d’un écran à travers une loupe. On voit distinctement sur l’agrandissement de droite les 3 sous-pixels rouge, vert et bleu
On peut par exemple voir sur la figure 4.2 la photographie de l’écran sur lequel ce texte est tapé: on voit nettement les alternances de pixels rouge, vert et bleu. Il faut cependant bien se rappeler que il ne s’agit que d’une reconstruction de la couleur qui ne sera valable que pour l’être humain. N’importe quel autre être vivant ayant des sensibilités différentes pour ses cônes et qui regarderait un écran de télévision ne verrait absolument pas une représentation fidèle des couleurs. Les dispositifs de représentation humains sont construits par et pour l’homme et ne représentent en aucun cas la radiation lumineuse réelle émise par un objet. Un spectromètre qui mesurerait le spectre de la lumière blanche en extérieur et la couleur “blanche” émise par un poste de télévision verrait d’une part un spectre continu et d’autre part trois pics situés sur les trois fréquences des primaires de l’écran.
On peut voir sur la figure 4.3 (partie gauche) les puissances à émettre par chacune des trois primaires pour reconstruire une couleur donnée. On se rend compte que la primaire rouge choisie à 700nm est extrêmement peu efficace et qu’il faut donc l’émettre dans une proportion beaucoup plus importante que les autres. Ces trois primaires sont importantes historiquement: elles ont été choisies pour les premières expérimentations au début du XXème siècle par Guild et Wright et sont les primaires du premier modèle colorimétrique, dit modèle CIE-1931 ou parfois RGB-1931; mais elles sont totalement inefficaces sur le plan pratique, et ne sont jamais utilisées.
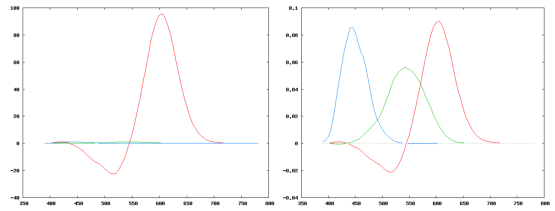
Figure 4.3: Puissances des trois primaires nécessaires à l’émission équivalente d’une couleur (brute à gauche, normalisée à droite)
On voit à droite la représentation normalisée des trois primaires. Il faut noter que pour générer une couleur équivalente à, par exemple, une radiation à 525nm, il faudrait générer une certaine quantité de rouge négatif, ce qui est évidemment impossible.
Les modèles colorimétriques définissent également la luminance, qui est la quantité supposée représenter l’intensité lumineuse ressentie. Pour le modèle RGB-1931, si (r,g,b) représentent les coordonnées d’un signal donné, la luminance est égale à:
| L(r,g,b) = 0.17697 r + 0.81240 g + 0.01063 b |
On appelle généralement chrominance les valeurs rr, g et b pour les distinguer de la luminance. Le codage d’information dit en composantes ou composite ou YCbCr code l’information lumineuse sous la forme (L,r,b), l’information concernant le vert se retrouvant en inversant l’équation de la luminance.
Les imprimantes quant à elles travaillent comme les peintres en synthèse soustractive. Elles utilisent des pigments qui absorbent certaines parties du spectre, d’où le nom de synthèse soustractive. La construction d’une couleur est absolument identique à celle du modèle additif, mais avec des primaires qui sont choisis comme les complémentaires des primaires du modèle additif. Le modèle additif est généralement dénommé modèle RGB pour Red (Rouge), Green (Vert) et Blue (Bleu), et le modèle soustractif modèle CMY, pour Cyan, Magenta, Yellow (jaune) (voir figure 4.4).
On voit donc que le modèle trichromique est central dans la compréhension de la vision de la couleur, et qu’il a été validé expérimentalement. Nous allons maintenant en voir les fondements génétiques, et comment ils expliquent certaines anomalies de la vision humaine.
Pour plus de détails, on peut se reporter ici.
On appelle caractère héréditaire l’ensemble des caractères d’un individu transmis par le père et la mère par les gènes issus des chromosomes. Ainsi, plusieurs membres d’une même famille présentent les mêmes caractères, comme nous le montre les arbres généalogique d’une famille.
Cette partie sur l’hérédité nous permet de démontrer comment l’enfant peut posséder des caractères communs à ceux de ses parents afin d’expliquer comment le daltonisme se retrouve dans une même famille.
Les caractères héréditaire se transmettent donc, de la manière suivante: un embryon est tout d’abord formé suite à la fécondation d’une ovule de la mère et d’un spermatozoïde du père. Chacune de ces deux cellules reproductrices contiennent alors un patrimoine génétique, localisé dans l’ADN, sur les chromosomes. Chacune de ces cellules reproductrice ou gamètes possèdent alors 23 chromosomes à un chromatides. L’ovule aura donc forcément un chromosome X tandis que le spermatozoïde aura soit un chromosome X, soit un Y. C’est donc le spermatozoïde qui déterminera le sexe du futur bébé, qui sera une fille si elle possède 2 chromosomes X et garçon s’il possède un chromosome X et un Y. La fécondation de ces 2 gamètes mâle et femelle aboutie à la formation d’ une cellule-œuf qui possédera désormais 46 chromosomes, hérité pour moitié de son père et pour moitié de sa mère. L’enfant possédera donc des caractères hérités de son père et de sa mère.
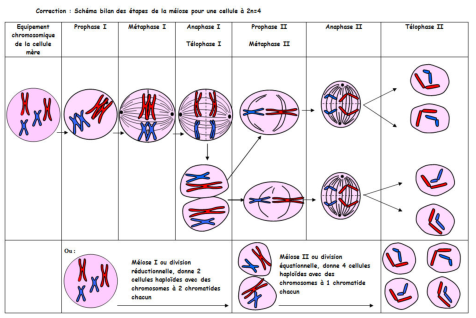
C’est lors de la méiose (voir figure 5.1), constituée de deux divisions cellulaire successive que vont être formées les gamètes. Ainsi, à l’issue de la méiose, une cellule originale dite cellule mère aura donné naissance à quatre cellules haploïdes (une cellule qui ne comporte qu’un seul exemplaire de chaque chromosome) dont le matériel génétique aura été recombiné de manière unique.
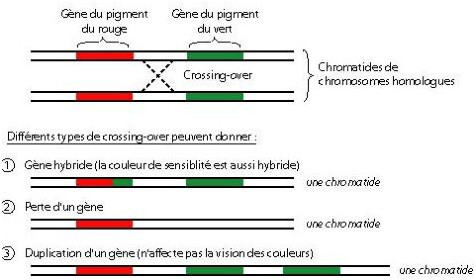
En effet, lors de la première division cellulaire de la méiose, les chromosomes vont migrer au centre de la cellule et chaque chromosome d’une même paire vont se réunir à coté de leur chromosome homologue (chromosome appartenant à la même paire, de même taille, possédant les mêmes gènes mais pas obligatoirement les mêmes allèles) au niveau de la plaque équatoriale. Les chromatides des paires de chromosomes homologues formeront ainsi des tétrades de chromatides. Au niveau de ces tétrades se produisent des phénomènes nommés crossing-over (ou enjambement génétique) qui correspond à un croisement puis un échange de certains segments d’ADN au niveau des chromosomes homologues, ce qui permet un brassage génétique (recombinaison génétique au sein d’une population entière et sur plusieurs générations). Des chromosomes recombinés se sont alors formés. Après ces échanges, les chromosomes homologues se séparent, un chromosome de chaque paire migre vers un des pôles de la cellule, et la première division cellulaire se termine. On a ainsi obtenu 4 cellules haploïdes avec des chromosomes à 1 chromatide chacun.
La seconde division cellulaire de la méiose est similaire à celle de la mitose sauf qu’elle concernes des cellules haploïdes (lorsque les chromosomes de la cellule sont chacun en un seul exemplaire, opposé à diploïde). A la fin de la métaphase, les centromères se fissurent et les 2 chromatides de chaque chromosome migrent vers les 2 pôles de la cellule: la division est alors terminée.
Ainsi, il y a donc eu un double brassage génétique au cours de la méiose:
Ces nombreuses recombinaisons permettent d’obtenir des gamètes uniques, ce qui explique en partie pourquoi chaque individu possède un patrimoine génétique spécifique, unique.
Il existe plusieurs modes de transmission des caractéristiques génétiques, suivant que le gène transmis se trouve sur un chromosome non sexué (autosome) ou sexué.
AA Aa aa AA AA: 100% AA: 50% Aa: 50% Aa: 100% Aa AA: 50% Aa: 50% AA: 25% aa: 25% Aa: 50% Aa: 50% aa: 50% aa Aa: 100% Aa: 50% aa: 50% aa: 100%
Pour une fille, la transmission est proche de celle faite avec un chromosome autosome. En revanche, pour un garçon, c’est uniquement la mère qui transmet aléatoirement un de ses gènes à son fils. S’il reçoit le gène muté a il présentera systématiquement le phénotype muté, que a soit récessif ou dominant.
AA Aa aa A AA: 100% AA: 50% Aa: 50% Aa: 100% a Aa: 100% Aa: 50% aa: 50% aa: 100% A: 100% A: 50% a: 50% a: 100%
Tableau 5.2: Gène porté par le chromosome X, transmission pour une fille en haut, puis pour un garçon en bas
Les maladies ou les défauts génétiques transmis par le chromosome X sont presque toujours majoritairement récessifs à quelques rares exceptions près (comme le syndrome de Rett). Dans ces très rares cas, le défaut se manifeste majoritairement de novo, c’est à dire qu’il se manifeste à la suite d’une mutation spontanée du génome alors que les deux parents sont chromosomiquement normaux. Généralement, seules les filles hétérozygotes survivent; les garçons (portant seulement le gène muté) ou les filles homozygotes pour le gène muté sont majoritairement non viables.
L’identification des gènes responsables du codage des opsines a commencé à partir des années 80 avec le développement des techniques de séquençage de l’ADN. Elles se sont poursuivi pendant les années 90 et les années 2000, pour en arriver aujourd’hui à une taxonomie à peu près complète qui permet de tracer comment au cours de l’évolution ces différents gènes ont évolué et se sont modifiés. On sait aujourd’hui que les opsines se sont différenciés au cours de l’évolution à partir de deux gènes originaux chez les vertébrés (voir figure 5.2). Ces deux gènes ont donné d’une part (pour le groupe L) les gènes codant les cônes sensibles aux rouges ou aux verts proches du rouge.
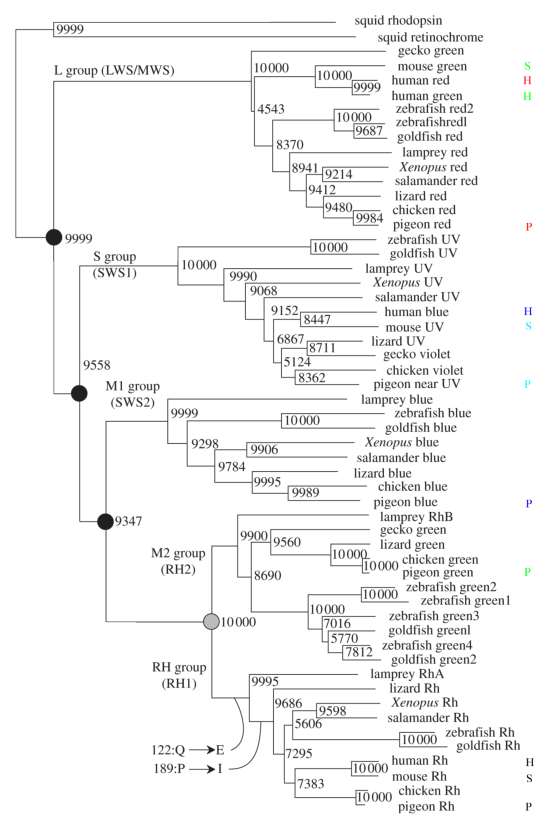
Figure 5.2: Évolutions des opsines. On a marqué les opsines humaines (H), celles de la souris (S) et du pigeon (P) (extrait de [SM09])
Le second gène s’est lui-même différencié en quatre groupes: SWS1 qui code les cônes bleus proches des UV et les cônes UV, SWS2 qui code les bleus proches du vert et les verts proches du bleu, RH2 qui code strictement le vert en luminosité plutôt faible, et RH1 qui code exclusivement la rhodopsine des bâtonnets: il y a donc quatre familles de gènes codant les photopsines et une famille codant la rhodopsine.
Comme on le voit sur la figure 5.3, les oiseaux ont conservé les quatre types de cônes liés aux quatre gènes principaux. Ces cônes ont une couverture spectrale de bien meilleure qualité que celle de l’homme, puisqu’elle couvre un spectre bien plus étendu, avec un recouvrement très faible. Les oiseaux sont donc susceptibles de discerner des “couleurs” qui nous sont inaccessibles, et tout autant impossible à comprendre que la différence entre le vert et le rouge pour un daltonien.
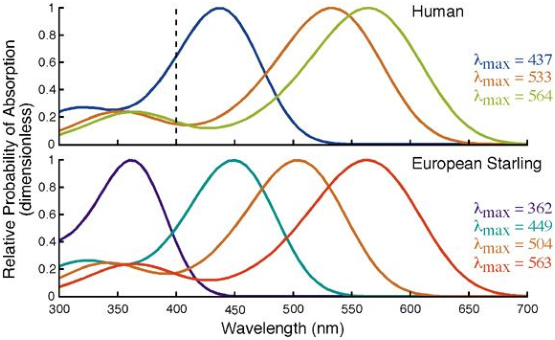
Figure 5.3: Sensibilité spectrale comparé des cônes des étourneaux et des humains (extrait de [Gol07])
La plupart des mammifères ont perdu deux (SWS2 et RH2) des quatre gènes des ancêtres communs lors de l’évolution des espèces. La raison semble être que les premiers mammifères étaient exclusivement nocturnes, et que l’évolution a donc favorisé le développement des bâtonnets dans l’œil au détriment des cônes.
Aujourd’hui, la quasi-totalité des mammifères sont donc dichromates, certains même, comme les raton-laveurs, sont totalement incapables de percevoir la couleur, car ils ne possèdent plus qu’un seul type de cône (ils ont également perdus le gène SWS1). C’est aussi le cas de certains lémuriens, qui sont, en terme d’évolution naturelle, relativement proches de l’être humain.
La version trichromique n’est réapparu que chez les primates (dont l’homme), à travers deux mécanismes différents (voir figure 5.4).
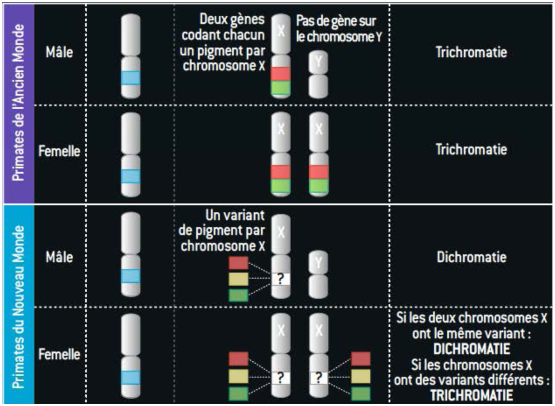
Figure 5.4: Réapparition de la trichromie chez les primates (extrait de [JN10])
Si le gène SWS1 se trouve sur un chromosome homologue (le chromosome 7), le gène MLWS se trouve sur le chromosome X. Au cours de l’évolution, des allèles différents sont apparus par mutation sur ce gène.
Chez les primates (homme, chimpanzé, gorille, etc) dits de l’ancien monde (Afrique, Europe, Asie), le gène s’est dupliqué et la forme majoritaire qui s’est fixé est celle de deux gènes très proches, à la fois spatialement (ils sont côte à côte) et en terme de séquence ADN. Ces deux gènes codent deux opsines qui sont aussi très proches (en raison de la quasi-similitude des séquences génétiques), ce qui explique que chez ces primates les sensibilités spectrales des cônes L et M se recouvrent largement (il est aussi fréquent de rencontrer plusieurs copies du gène encodant l’opsine verte sur le chromosome X (généralement deux)).
Tous ces résultats sont confirmés par la génétique moléculaire qui montre que les gènes LW et MW sont identiques à 96%, alors qu’ils n’ont qu’environ 40% de nucléotides en commun avec les gènes codant l’opsine bleu et la rhodopsine des bâtonnets. L’opsine rouge et verte ne se distinguent que par 15 acides aminés, dont 3 seulement (en position 180, 277, et 285) semblent avoir une influence majeure sur la longueur d’onde capturée.
Chez les primates dits du nouveau monde (Amérique), il existe plusieurs allèles différents (principalement trois), mais une seule copie du gène. Seules les femelles possédant deux allèles différents sur les deux chromosomes X sont trichromates en raison du mosaïcisme lié à l’inactivation aléatoire d’un chromosome X. Tous les mâles ainsi que les femelles ayant des allèles identiques sont dichromates.
La réapparition de la différenciation rouge–vert chez les primates semble lier à l’avantage évolutif que leur donne la capacité de mieux distinguer le vert du rouge, ce qui était particulièrement important pour des populations de cueilleurs qui devaient distinguer les baies mures généralement rouges, du feuillage et des baies non mures généralement vertes (voir figure 5.5).

Figure 5.5: Une baie mûre vue dans son environnement par un trichromate (à gauche) et un dichromate (à droite)
Les conséquences pour l’homme de ces structures chromosomiques sont nombreuses:
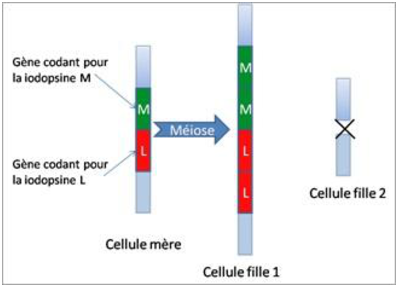
L’autre chromosome X, en revanche, possède le gène ( L ou M) en double exemplaire, et l’homme qui en hérite à une vision colorée normale: en effet, le gène supplémentaire n’a pas d’effet.
On peut voir sur la figure 5.6 un résumé de diverses possibilités liés au crossing over.
C’est ce phénomène qui explique d’une part la grande fréquence de certaines formes de daltonisme, et d’autre part qu’il existe aussi chez l’homme plusieurs formes alléliques pour ces deux gènes.
Nous allons maintenant détailler les différentes formes de défauts de la vision liés aux gènes encodant les opsines, généralement désignés sous le nom de daltonisme.
Le terme de daltonisme est relativement malheureux. Les anglo-saxons ne l’emploient pas, préférant le terme color-blindness, que l’on pourrait traduire par “aveugle aux couleurs”. Le terme vient du nom du chimiste britannique John Dalton qui était atteint de cette condition, et fut un des premiers à en rechercher scientifiquement les causes. Le terme scientifique correct est dyschromatopsie.
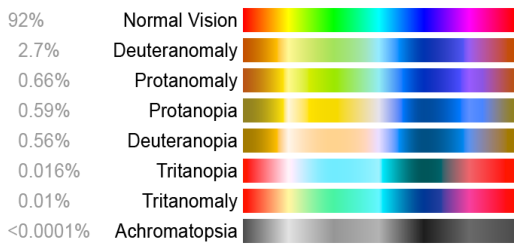
Nous allons voir qu’il existe plusieurs types de “daltonisme”. Tous ceux qui sont causés par les gènes codant les opsines rouges et vertes se transmettent par le chromosome X de façon récessive. Cela signifie que si une proportion p de la population masculine est affectée, la proportion de femmes affectées ne sera que de p2. La pénétration du daltonisme dans la population varie suivant les pays. En France elle est de l’ordre de 8% pour les hommes.
On voit sur la figure 5.7 une représentation des différents spectres accessibles aux genres atteints par cette condition.
John Dalton était atteint de deutéranopie comme l’a confirmé une analyse ADN d’un de ses globes oculaires effectué en 1995. Les autres formes de déficience des couleurs ne sont des daltonismes que par abus de langage.
Nous avons rapidement étudié dans la section précédente les gènes encodant les différentes opsines de l’œil. Mais la vision de la couleur est aussi dépendante du bon fonctionnement des différents composants des cônes, et il existe un certain nombre de mutations “relativement” fréquentes qui entraîne une achromatopsie totale, puisque aucun des cônes n’est susceptible de fonctionner correctement.
Voici une liste des gènes qui sont connus aujourd’hui comme intervenant dans l’achromatopsie totale. Certaines mutations intervenant sur ces gènes ne causent que des achromatopsies partielles, car les protéines codées restent partiellement actives. Ces transmissions sont toutes autosomales récessives.
On voit ici que l’on n’explique actuellement qu’environ 80% des cas d’achromatopsie totale. On recherche encore les autres causes génétiques de cette condition. D’autres pistes sont suivies, dont des anomalies qui seraient liées à des gènes présents sur le chromosome 14.
Il faut noter qu’il existe des formes d’achromatopsie congénitale qui s’accompagne de désordres beaucoup plus importants au niveau de la vision, comme la perte de la reconnaissance des formes, et bien d’autres déficits graves qui peuvent être liés à des erreurs génétiques sur des gènes encodant des protéines indispensables au bon fonctionnement du cortex cérébral ou du système nerveux. Nous n’en parlerons pas ici.
Dans les précédents chapitres, nous avons vu que de nombreuses caractéristiques de la vision humaine et de ses anomalies pouvaient s’expliquer par le modèle trichromique et la génétique qui le sous-tend.
Pourtant, comme nous l’avons vu dans l’introduction, ce modèle n’explique pas certains paradoxes de la vision humaine, qui trouvent leur origine dans le fonctionnement du traitement cérébral de l’information lumineuse. C’est ce que nous allons voir maintenant.
Le traitement par la rétine et le cerveau de l’information fournie par les différents récepteurs que sont les cônes et les bâtonnets est incroyablement complexe. La science évolue énormément dans ce domaine depuis le milieu de XXème siècle, avec des revirements fréquents. Par exemple, on croyait au début des années 80 que l’aire V4 du cortex visuel était chargé du traitement de la couleur alors que l’on pense aujourd’hui que ce n’est pas du tout le cas. Le livre de Semir Zeki ([Zek93]) qui faisait autorité au début des années 90 est considéré comme dépassé, et même inexact par endroits.
Dans un article faisant le panorama de la recherche dans ce domaine en 2011, Robert Shapley ([SH11]) écrit:
The main point of our review of visual neuroscience over the past 25 years is that color is not separate and apart, but rather that color and form and motion are inextricably linked as properties of objects in visual perception and in the visual cortex..Space and color are not distinct elements but, rather, are interdependent aspects of a unitary process of perceptual organization.
On voit bien sur la figure 6.1 que les perceptions des formes et des couleurs sont fortement liées, comme ce type d’illusions visuelles le montrent.

Figure 6.1: En regardant l’image du haut, on ne distingue rien, alors que la carte de la méditerranée apparaît clairement sur celle du bas. Les perceptions de couleur et de forme sont dépendantes (extrait de [WPS07])
Les modèles modernes du traitement de l’information de couleur par le cerveau sont trop compliqués et évoluent trop vite pour que nous les présentions ici. Nous allons nous contenter de présenter les faits qui semblent acquis et nous limiter au traitement fait dans la rétine, avec quelques éléments de ce que l’on croit savoir concernant le corps géniculé latéral (CGL) et l’aire V1 du cortex visuel.
De façon schématique, le traitement de l’information visuelle commence dans la rétine.
Il existe ensuite deux voies principales: la voie non visuelle et la voie visuelle. La voie non-visuelle est dirigée vers le noyau suprachiasmatique. Elle reçoit des informations des cônes et des bâtonnets, mais aussi des cellules ganglionnaires photosensibles. Cette voie est principalement impliquée dans la détermination des rythmes circadiens; le noyau suprachiasmatique est en particulier impliqué dans la détermination des cycles du sommeil, mais aussi dans la régulation de la température corporelle. Il semble que les cellules ganglionnaires photosensibles n’interviennent pas dans la voie visuelle, même si cela ne peut être complètement exclu.
L’information sur la voie visuelle est véhiculée par les nerfs optiques vers le chiasma optique où les deux nerfs optiques se rassemblent. A ce niveau, l’ensemble des axones venant des deux hémi-rétines gauches sont dirigés vers le corps géniculé (ou genouillé) gauche de l’encéphale, et l’ensemble des deux hémi-rétines droites vers le corps géniculé droit.
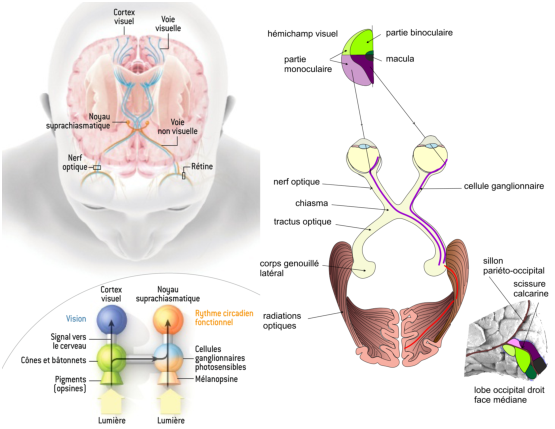
Figure 6.2: Voies visuelles et non visuelles (extrait de [Pro12])
De là, l’information est dirigée vers le cortex visuel (voir figure 6.2).
Nous allons maintenant détailler le fonctionnement de la voie visuelle dans les sections suivantes. Il faut cependant bien se rappeler encore une fois que les modèles de traitement de l’information visuelle dans le cerveau sont en permanente évolution, et que nous n’en présentons que la partie qui semble à peu près “sûre” aujourd’hui.
La rétine a une fonction importante à remplir, celle de compresser l’information en provenance des cônes et des bâtonnets pour qu’elle puisse être transmise au cerveau via le nerf optique. En effet, le nerf optique ne comporte qu’un à deux millions de fibres nerveuses, alors que la rétine comprend entre 100 et 200 millions de récepteurs.
La structure de la rétine est elle-même très compliquée, et le mode de fonctionnement de certains composants ne sont pas totalement connus. La rétine comprend (voir figure 6.3):
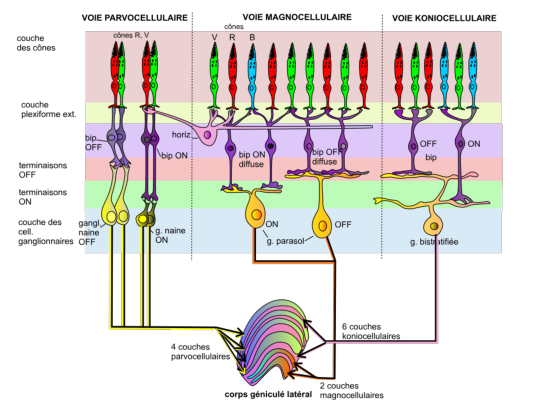
On voir sur la figure 6.4 les différentes voies de transmission de l’information vers le CGL.
Il existe principalement trois voies permettant de transférer l’information de la rétine vers le corps géniculé latéral:
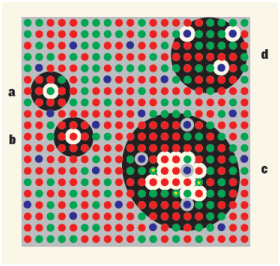
Figure 6.5: Différents types de cellules ganglionnaires(extrait de [DB10])
On voit sur la figure 6.5 le fonctionnement détaillé de différentes cellules ganglionnaires impliquées dans le transfert des informations chromatiques:
On voit bien que le mécanisme classiquement décrit sous la forme d’un cône excitateur au centre opposée à des cônes périphériques inhibiteurs n’est valable que pour les cellules centrales de la fovéa.
On voit aussi que l’information de chrominance transmise au cerveau est donc constitué d’un canal V−R et d’un canal B−(V+R).
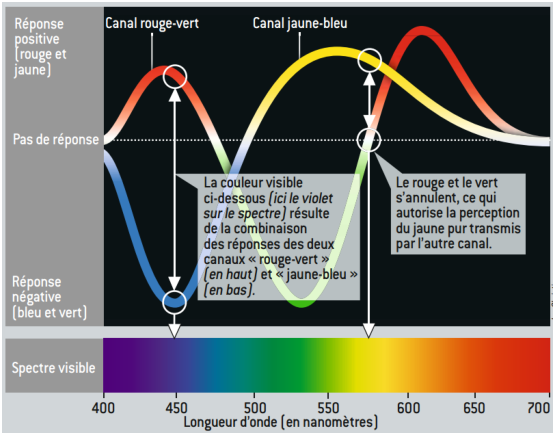
Figure 6.6: Le mécanisme d’opposition explique pourquoi certaines couleurs sont “visibles” et d’autres sont “interdites” (extrait de [BT10])
C’est le codage de l’information sous cette forme qui explique comment le cerveau va percevoir les radiations, comme cela est expliqué sur la figure 6.6 où l’on peut voir comment le spectre lumineux est encodé par le double canal R−V et B−(V+R).
C’est en particulier pour cette raison que l’on est capable de percevoir une continuum régulier de couleurs entre le bleu et le vert, alors que le rouge-vert ou le vert-rouge n’existe pas.
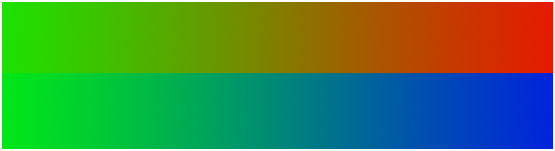
Figure 6.7: Le vert se dissout dans le bleu, alors que l’on distingue clairement le jaune et l’orange lors du passage du vert au rouge
On le voit ainsi sur la figure 6.7. On passe du bleu au vert en passant par du bleu-vert, alors que le passage du vert au rouge montre clairement le jaune et l’orange.
Au niveau du CGL, on sait que les 3 voies P, M et K décrites ci-dessus restent fonctionnellement séparées et vont alimenter séparément la partie V1 du cortex visuel. On sait également que le CGL comprend des cellules à opposition simple comme dans la rétine. On pense que le CGL est également impliquée dans l’augmentation des contrastes et qu’il maintient, et même renforce la séparation R−V et B−(R+V), mais son rôle n’est pas complètement clair. Le CGL transmet ensuite l’information à la zone V1 du cortex visuel.
Le fonctionnement de la zone V1 du cortex visuel est aujourd’hui encore mal comprise. On sait, à partir de tests faits par imagerie à résonance magnétique (IRM) que les cellules de V1 ont des propriétés complexes: certaines d’entre elles sont par exemple spécialisées pour répondre à des stimuli orientés spatialement. Cette zone a aussi un rôle important dans le traitement des informations de couleurs à travers une classe particulière de cellule, les cellules à double opposition (“double opponent cell”).
Nous avons vu dans le paragraphe précédent que le codage de l’information qui est fait dans la rétine passait par des cellules à simple opposition: rouge central moins vert périphérique, ou vert central moins rouge périphérique, etc... Les cellules à double opposition ont été découvertes très récemment chez l’homme (moins de dix ans), et elles fonctionnent sur un modèle différent (voir figure 6.8).
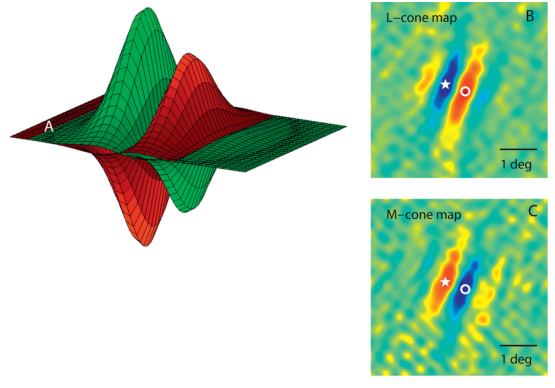
Figure 6.8: Réponse d’une cellule en double opposition de l’aire V1 du cortex visuel (extrait de [SH11]).
Pour une cellule en double opposition, la sortie positive est maximale quand la zone de gauche reçoit un signal vert et la zone de droite un signal rouge. Elle sera également très forte mais dans le sens négatif si la zone de gauche reçoit un signal rouge et la zone de droite un signal vert. Si les deux zones reçoivent des signaux uniformément rouges ou verts, la sortie est nulle.
Ces cellules sont généralement raccordées à travers le CGL à des zones de taille relativement grandes de photorécepteurs. Elles sont donc particulièrement adaptées à reconnaître des différences de chrominance et ce quel que soit l’éclairage ambiant.
Les études les plus récentes semblent montrer que ce sont ces cellules qui sont effectivement responsables du phénomène connu sous le nom de consistance de la couleur, dont nous avions déjà parlé dans notre introduction.
La consistance de la couleur est le phénomène qui permet à la vision humaine de conserver, quelle que soit la lumière ambiante, la même couleur à un objet (voir figure 6.9).
Ce phénomène a été étudié extensivement dans les années 70 par Edwin Land (le fondateur de Polaroïd), qui a développé une théorie (la théorie retinex [Lan78]) qui a servi et sert encore comme algorithme pour équilibrer la balance des blancs dans les appareils photos numériques; même si les résultats obtenus en biologie aujourd’hui infirment partiellement la théorie de Land, ces algorithmes sont toujours utilisés.
L’aire V1 du cortex visuel semble aussi impliqué dans un autre phénomène connu sous le nom d’assimilation de couleur, même si l’on n’est pas complètement certain que ce soit au niveau de V1 que ce phénomène se produise.
Les mécanismes impliqués semblent plus complexes et encore mal compris, ce qui montre bien comme il est encore aujourd’hui difficile d’expliquer la totalité des mécanismes de la vision humaine, même sur un exemple aussi simple que celui présenté ci-dessous.
Il s’agit en effet d’un phénomène qui n’est pas explicable par les mécanismes que nous avons décrit jusque là et dont on voit un exemple sur l’illusion visuelle qui est présentée sur la figure 6.10.

Figure 6.10: Exemple d’assimilation de couleur (extrait de [Con09]).
Le “C”, le “l” et le “r” de “Color” sont tous de la même couleur turquoise . Le “C” et le “r” sont traversés de bandes violettes, alors que le “l” est traversé par des bandes jaunes. Associé avec du jaune, le turquoise parait plus jaune, ce qui est incohérent avec la théorie qui voudrait que cela renforce le contraste avec le bleu, et qu’il devrait donc paraître davantage bleu et non davantage jaune.
Le même phénomène se produit avec les deux “o” qui sont tous les deux de la même couleur orange, mais le “o” traversé de bandes jaunes parait plus jaune, ce qui est là aussi incohérent.
Nous avons essayé de montrer que la vision de la couleur est profondément dépendante du traitement de l’information par le cerveau. On peut également le montrer en constatant que ce traitement n’est pas le même pour l’ensemble du règne animal, et que cela entraîne des différences profondes dans la perception de la couleur.
Ainsi, Décrire le processus de la vision ne peut se faire que si l’on comprend le mécanisme de traitement de l’information; connaître le nombre de photorécepteurs et le spectre qu’ils couvrent n’est qu’une partie très limitée du problème.
On sait par exemple depuis peu ([Jac14]) que si le système de traitement de l’information est probablement à peu près le même chez tous les mammifères, certains animaux ont des systèmes de traitement plus primitifs, mais peut-être plus efficaces en terme de consommation d’énergie.
On sait par exemple que les crevettes-mantes disposent de 12 types de photorécepteurs différents (voir figure 6.11). Pourtant elles sont moins capables de distinguer des couleurs proches que l’être humain. Là où nous sommes capables de distinguer des nuances de l’ordre du nanomètre, les crevettes-mantes semblent incapables de distinguer des nuances en dessous de 20nm d’écart.
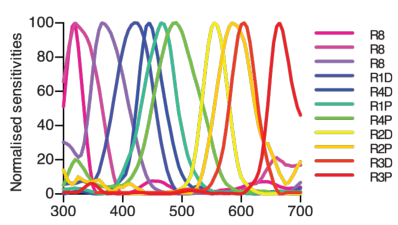
Figure 6.11: Les 12 types de photorécepteurs des crevettes-mantes (extrait de [THCM14])
Il s’agit là d’une preuve supplémentaire de l’interdépendance entre les différents composants du système de la vision.
Il existe de nombreux troubles cérébraux qui se manifestent sous de multiples formes. Certains ne modifient pas directement la capacité à distinguer les couleurs, mais modifient d’autres caractéristiques plus subtiles. Il existe par exemple des patients présentant les formes suivantes:
Mais les défauts de la vision liés aux traumatismes cérébraux entraînent généralement une achromatopsie cérébrale, c’est à dire la perte totale ou partielle de la vision des couleurs liée à un dysfonctionnement cérébral. Il faut bien distinguer l’achromatopsie cérébrale de l’achromatopsie dont nous avons parlé au chapitre précédant, et qui est du à des mutations des gènes codant des composants fondamentaux des cônes.
L’achromatopsie cérébrale est la conséquence de dommages au cerveau liés en général à une privation d’oxygène du cortex visuel à la suite d’un accident vasculaire cérébral, ou d’une ischémie (noyade, incendie,…), ou à des maladies telles que l’herpès, l’encéphalite ou la méningite. Certains problèmes congénitaux peuvent également entraîner une achromatopsie.
L’achromatopsie cérébrale se manifeste sous des formes proches, bien que subtilement différentes. Elle s’accompagne souvent de l’incapacité de reconnaître les visages (prosopagnosia), ou parfois les formes, ce qui tendrait à prouver que ces traitements sont liés et renforce une fois de plus l’idée que les traitements visuels de l’information ne sont pas séparés comme on le pensait à une époque.
Pour tester ces hypothèses, on a de plus en plus recours à l’imagerie par résonance magnétique. Il s’agit de tester un certain nombre de patients et d’évaluer les flots d’oxygène dans le cerveau, l’oxygène comportant de l’hémoglobine qui contient du fer, et qui est donc détectable par magnétisme.
On teste alors des patients souffrant d’achromatopsie et des patients souffrant de prosopagnosia, et l’on essaie de voir quelles zones du cerveau ont été détériorés par un accident ischémique en les comparant avec des patients ayant une vision normale de la couleur et de la reconnaissance des formes.
On regarde ensuite comment les différentes images se recouvrent et on essaie d’en déduire les zones impliquées dans ces différents phénomènes; le but est de rechercher des zones de superposition.
En fait, si l’on distingue bien des zones communes entre l’achromatopsie et la prosopagnosia, on se rend aussi compte de la difficulté de distinguer clairement une seule zone impliquée dans les deux processus.
Comme on le voit sur la figure 6.12, les résultats ne sont pas strictement localisés. Il faut aussi dire que certains chercheurs mettent aujourd’hui partiellement en cause les résultats obtenus par IRM, en raison de l’opacité des logiciels assurant le traitement des images et de la fiabilité relative des IRM à détecter en temps réel des processus transitoires.
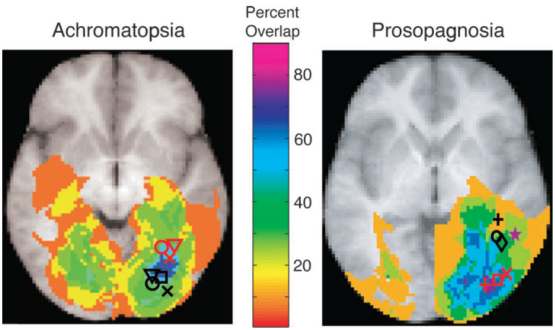
Figure 6.12: Superposition de lésions par des moyens d’imagerie médicale. A gauche, achromatopsie, à droite prosopagnosia. Les symboles noirs ou rouges donnent des indications sur la coordonnée z qui est invisible sur des scans en 2D (extrait de [BE06]).
Les patients souffrant d’achromatopsie cérébrale sont incapables de distinguer les couleurs: un des tests classiques pour les dépister est l’utilisation des tests de Farnsworth-Munsell et d’Ishihara (nous reviendrons plus en détail sur le test d’Ishihara dans le chapitre 7. Il faut noter que ces deux tests, qui sont aussi efficaces pour détecter les défauts de vision de la couleur liés aux opsines de l’oeil ne le sont pas dans le cadre de l’achromatopsie cérébrale.
On peut voir sur la figure 6.13 le principe de fonctionnement de ces deux tests.
Les patients sont incapables d’effectuer correctement le test de Farnsworth-Munsell alors qu’un certain nombre d’entre eux arrivent à effectuer le test d’Ishihara, à condition que les figures leur soient présentées à une distance relativement grande.
L’éloignement de la figure fait disparaître la vision des détails de l’image, et ne met plus en cause que la vision du contraste de la couleur, qui est fondamentale dans le test d’Ishihara.
On pense ainsi que la zone lésée ne met pas en cause les zones de V1 qui sont responsables de ce mécanisme (les cellules à double opposition dont nous avons parlées dans la section précédente), ce qui expliquerait que, bien que la vision des couleurs soit perdue, le contraste visuel continue à fonctionner correctement, même si les patients n’en sont pas nécessairement complètement conscients. Il s’agit là encore d’un argument de plus dans l’hypothèse du traitement complexe et multi-zonal de l’information de la couleur.
Un autre exemple intéressant est celui d’un patient souffrant d’achromatopsie complète à la suite d’un herpès cérébral. Ce cas, qui a été largement étudié dans la littérature médicale, a donné lieu à des expériences particulièrement frappantes. On peut en voir une sur la figure 6.14.

Figure 6.14: Deux phrases superposées, l’une en noir sur noir et l’autre en rouge sur vert. Une personne normale pourra lire la seconde mais pas la première. Un achromate “rétinal” ne pourra lire ni l’une ni l’autre. Certains achromates cérébraux peuvent lire la seconde comme s’ils percevaient la couleur, bien qu’ils soient incapables de distinguer la moindre différence de couleur. (extrait de [KHC04])
Cet exemple montre bien que la notion de contraste de couleur est conservée alors que la capacité à distinguer les couleurs a disparu, ce qui s’explique à nouveau par un fonctionnement correct de la partie de V1 qui assure la distinction du contraste de couleur, mais une perte de fonctions d’ordre supérieur, non encore identifiée.
Il reste encore beaucoup de chose à découvrir sur le fonctionnement du cerveau pour l’appréhension de la couleur. Certaines des découvertes récentes ont permis de comprendre certains déficits, mais elles ont surtout montré que le modèle est beaucoup plus complexe et interdépendant qu’on ne le pensait il y a une vingtaine d’années.
Le dépistage du daltonisme est pratiqué systématiquement dans la majorité des pays développés. Les tests pour détecter le daltonisme sont très anciens, et remontent à l’origine de l’étude de la perception des couleurs.
Le plus utilisé aujourd’hui est le test d’Ishihara. Le test chromatique d’Ishihara fut nommé d’après le nom de son inventeur, le docteur Shinobu Ishihara, professeur à l’université de Tokyo, qui publia pour la première fois ses tests en 1917.
Il est utilisé pour dépister les anomalies de la vision des couleurs. Il permet de détecter toutes les déficiences dyschromatiques sauf la tritanopie et la tritanomalie, qui sont très rares.
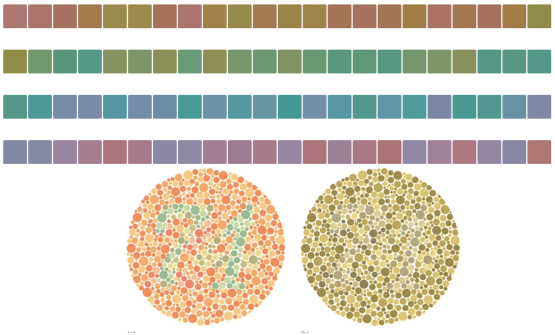
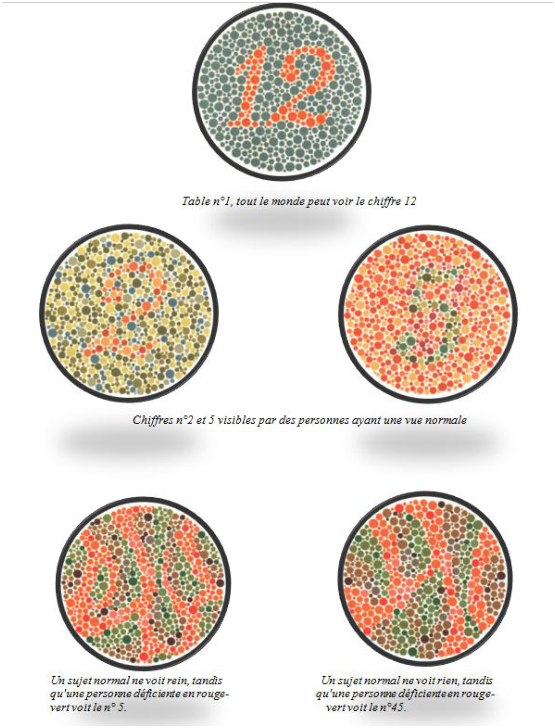
Le test se compose de trente-huit planches colorées. Sur chacune de ces planches, on a construit un cercle constitué de points de différentes tailles et de différentes couleurs, apparemment disposés de manière aléatoire. Les planches sont faites de façon à ce que quelqu’un disposant d’une vue normale soit capable de lire un nombre dont le contour est clairement délimité par des différences de couleurs.
Quelqu’un dont la vue présente certaines différences sera soit incapable de lire ces nombres, soit sera capable pour certaines planches de distinguer des nombres qui sont invisibles pour quelqu’un ayant une vision des couleurs normale.
On peut voir sur la figure 7.1 quelques exemples de ces planches.
http://le-daltonisme-origines.e-monsite.com/
http://fr.wikipedia.org/wiki/Daltonisme
http://www.daltoniens.fr/
http://www.doctissimo.fr/html/sante/bien_voir/sa_4944_daltonisme.htm
http://le.daltonisme.free.fr/
http://www.le.daltonisme.sitew.com/Le_daltonisme.E.htm#Le_daltonisme.E
http://www.bioinformatics.org/oeil-couleur/dossier/daltonisme.html
http://ophtasurf.free.fr/lavision.htm
http://www.podcastscience.fm/dossiers/2010/09/22/dossier-le-daltonisme-les-daltoniens/
http://stephane.chalamet.free.fr/html/images/memdalto.pdf
http://www.universalis.fr/encyclopedie/daltonisme/3-heredite-du-daltonisme/
http://tpedaltonisme09.jimdo.com/2%C3%A8me-partie-1/2-le-daltonisme-et-son-origine/
https://sites.google.com/site/llfems195/physiologie-de-l-oeil
http://www.svt-monde.org/spip.php?article35&artsuite=2
http://www.edition-sciences.com/
http://www.cnrs.fr/
http://www.bioinformatics.org/oeil-couleur/dossier/book.pdf
http://fr.wikipedia.org/wiki/%C5%92il
http://www.futura-sciences.com/magazines/sante/infos/dico/d/medecine-retine-4322/
http://lecerveau.mcgill.ca/
http://www.explic.com/20941-optique.htm
http://www.leglaucome.fr/
http://www.teteamodeler.com/sante/vue/vue6.asp
http://www.medecine-et-sante.com/anatomie/anatomieoeil.html
Ce document a été traduit de LATEX par HEVEA